Видільна система, екськреторная система, сукупність органів, що виводять з організму тварин і людини надлишок води, кінцеві продукти обміну, солі, а також отруйні речовини, що введені в організм або утворилися в нім. В простих легкорозчинні екськрети (аміак, сечовина) виводяться в довкілля шляхом дифузії (морські і паразитичні форми) або за допомогою скоротливих вакуолей, що виконують в основному функцію осморегуляції (в прісноводих форм). В нижчих водних багатоклітинних (губки, кишечнополостниє) і у малоактивних морських тварин (голкошкірі) продукти обміну, в основному аміак, дифундують через поверхню тіла і стінки порожнин, пов'язаних з довкіллям. Зокрема видільна функція у більшості безхребетних властива кишечнику.
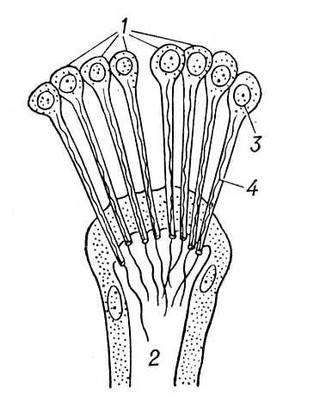
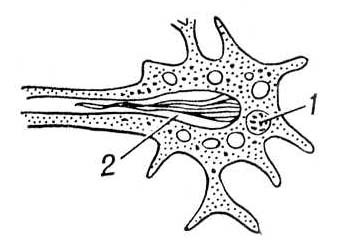
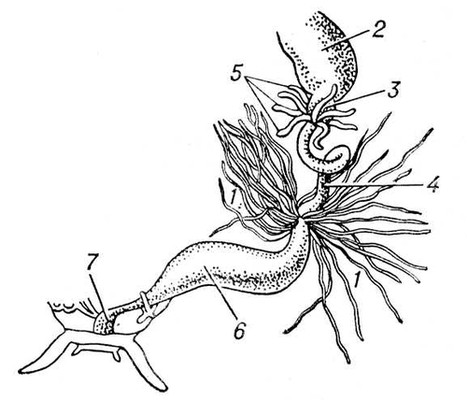
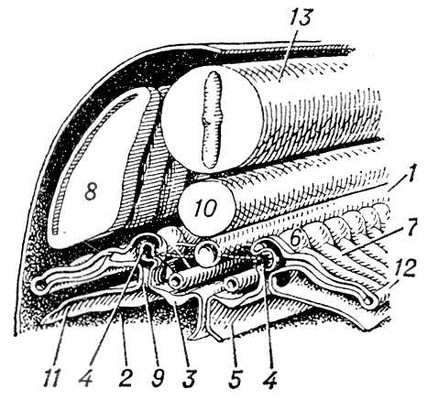
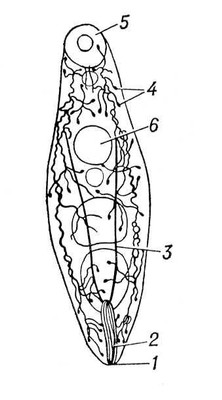
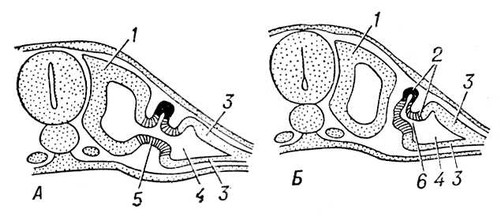
В процесі еволюції диференціюється спеціальна Ст с. У нижчих черв'яків (плоскі, гастротріхи, кинорінхи, коловертки, шкрябни, немертіни), а також в пріапулід і деяких аннелід і в личинок поліхет і молюсків видільні органи — протонефрідії — система трубочок, що відкриваються назовні порами, а що усередині сліпо закінчуються особливими порожнистими клітками, в яких знаходяться пучки довгих злиплих вій, що безперервно здійснюють коливальні рухи («миготливе полум'я») ( мал. 1 , 2 ). У кільчастих черв'яків сліпі кінці протонефрідіальних каналів закінчуються групами пляшковидних кліток (соленоцитов), у вузькій частині яких знаходиться по одному джгутику ( мал. 3 ). У більшості кільчастих черв'яків Ст с. — метанефрідії, тобто метамерний розташовані парні трубчасті ектодермальні органи, що відкриваються в целомічні мішки війковою воронкою, а назовні — деколи на поверхні наступного сегменту. Якщо до складу таких органів виділення входять і мезодермальниє елементи, то говорять про нефроміксій. Видільні органи, що розвиваються з мезодерми, — так звані целомодукти — є у молюсків: воронка відкривається в навколосерцеву порожнину (залишок целома ), зовнішній отвір — в мантійную порожнину. В ракоподібних видільні органи — звиті трубки, що починаються сліпим «целомічним» мішечком і що відкриваються назовні в підстави антен або максилл (звідси назви «антеннальниє» і «максилярні» залози) ( мал. 4 ). Схожі видільні органи в мечехвостов («коксальниє залози»), що відкриваються в підстави 5-ої пари ніг. Видільні органи, що відкриваються назовні, через які виводяться легко розчинні продукти обміну, типові для водних безхребетних.
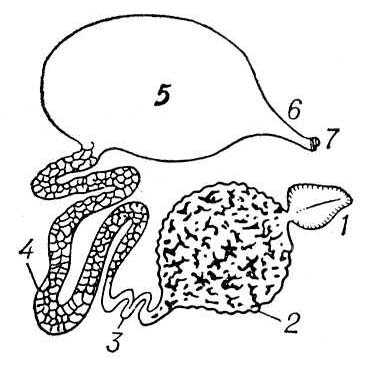
Проживання на суші, що вимагає економії витрати вологи, викликає зміну характеру катаболітів — легко розчинні аміак і сечовина замінюються важко розчинними гуаніном (в павукоподібних) або сечовою кислотою (у багатоніжок і комах). Видільні органи типа целомодуктів з наземних безхребетних мають лише вологолюбні форми — оніхофори і деякі павукоподібні (наприклад, сінокоси). Для наземних членистоногих характерний перехід видільної функції до стінок кишечника (стають інколи єдиним видільними органом, наприклад, в ложноськорпіонов) або частіше до мальпігиевим судинам — довгим сліпим виростам кишечника на кордоні середньої і задньої кишки, таким, що утворюється як вирости середньої (павукоподібні) або задньої (комахи, багатоніжки) кишки ( мал. 5 ). Фізіологічна перевага мальпігиевих судин (при проживанні в умовах дефіциту вологи) в тому, що через них легко випадні в осад продукти обміну (сечова кислота і ін.) виводяться з сечовою рідиною не назовні, а в задню кишку, де відбувається всмоктування води; зневоднені екськрети разом з неперетравленими залишками їжі виводяться через анальний отвір назовні, чим досягається збереження води в телі тварини.
Своєрідний тип виділення — відкладення переведених в труднорастворімую форму катаболітів (сечова кислота і ін.) в спеціальних клітках — «нирках накопичення» в нематод, в хлорагогенних клітках у дощових черв'яків, в проміжній тканині у павуків, в жировому телі у комах і багатоніжок, в печінкових виростах у стоног і т.п., що теж пов'язане з економією витрати води при екскреції. Схожі з цими видільними органами і екськретофори — клітки, нагромаджуючі екськрети і що викидають їх, наприклад, в просвіт кишечника (в ложноськорпіонов, у восьмипроменевих коралів і ін.).
Вторинний перехід наземних безхребетних до життя у воді, наприклад, личинок комах, привів до повернення екськретов до характерного для водних тварин типа (основний катаболіт — аміак) і до зростання осморегуляторной функції видільних органів (мальпігиеви судини і кишечник).
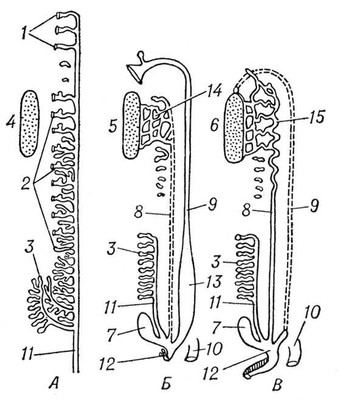
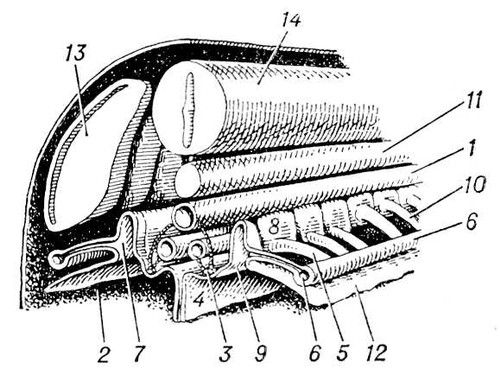
Серед хордових у покривників і асцидій органами виділення служать мішечки накопичення (у деяких покривників видільні елементи не виявлені). У ланцетника в зябрової області є близько 100 пар так званих нефроміксиев, які одним отвором відкриваються в околожаберную порожнину, а декількома іншими отворами (густо усадженими соленоцитамі) пов'язані з порожниною тіла. В хребетних органами виділення служать типові целомодукти, скупчення яких утворюють нирки . Від нирок відходять сечоводи, які впадають безпосередньо в клоаку або в сечовий міхур, що відкривається назовні сечовипускальним каналом. У еволюційному ряду хребетних, як і при індивідуальному розвитку вищих хребетних, спостерігається послідовна зміна трьох типів нирок. Першими виникають передні, або головні, нирки, звані також переднирками (див. Пронефрос ). Далі розвиваються первинні, середні, або тулубові, нирки — вольфови тіла (див. Мезонефрос ). Останніми з'являються вторинні, задні, або тазові, нирки (див. Метанефрос ). Целомодукти передбруньки і первинної бруньки, зазвичай по парі на сегмент, виникають з так званого нефротома. Частина нефротома, що відкривається у вторинну порожнину тіла, утворює миготливу воронку (нефростом). Проти нефростомов передбруньки в порожнині тіла зазвичай розвивається загальний для всіх воронок клубок кровоносних судин; через нього фільтруються з крові рідкі продукти виділення (водні розчини солей), які і поступають у воронки. Передбрунька, що має сегментарну будову і що тягнеться уздовж всього тулуба, зберігається в продовженні всього життя з круглоротих лише в бделлостоми. У міног залишки пронефроса, створюючі передній відділ бруньки, відкриваються в перикардіальну порожнину. В інших хребетних пронефрос служить лише личинковим органом. Він особливо добре розвинений в личинок більшості риб і земноводних. У всіх риб і земноводних в дорослому стані функціонують первинні нирки. Канальці їх розвиваються у ембріона схоже з канальцями передбруньки, відрізняючись лише тим, що відкриваються в готову пронефрічеський протоку, яка отримує з того часу назву первічнопочечного каналу ( мал. 7 і 8 ). У акулових риб цей канал розщеплюється з переднього кінця на дві протоки: вольфов канал службовець сечоводом первинної бруньки, і мюллеров канал, вступаючий в зв'язок із залишками пронефроса і що виконує у самок функцію яйцепровода. У самців земноводних вольфов канал знаходиться у зв'язку із статевою залозою і виконує одночасно функції і сечоводу, і семяпровода. Для первинної бруньки характерна наявність мальпігиевих телець . В амніот вторинна брунька замінює первинну бруньку, що функціонує в ембріональному періоді, яка у дорослих самок взагалі дегенеріруєт, а у самців несе функцію семяпроводящего апарату і називається придатком семенника, або епідидимісом. Ембріональний вторинна брунька утворюється із спеціального сечоводу, що зростає із заднього кінця вольфова каналу ( мал. 9 ). Цей сечовід має на кінці декілька кінцевих розгалужень, в які впадають ниркові канальці що виникають з нефрогенної тканини. Звиті канальці вторинної бруньки ніколи не мають воронок і починаються боуменової капсулою . У ссавців і людини за капсулою слідують проксимальниє звиті канальці, далі прямі низхідні і висхідні канальці, створюючі так звану петлю Генле, і, нарешті, дистальні звиті канальці і потім сполучні трубки, що впадають у вивідну протоку. Розширена частина сечоводу, звідки відходять його кінцеві розгалуження в бруньку, отримує назв.(назва) ниркової балії. Трьох типів нирок зв'язано між собою відомими переходами. Так, навіть в передбруньці можуть бути окремі клубочки, властиві первинній бруньці. У останній досить часто зникають воронки, і вона стає дуже схожою по будові на вторинну бруньку.
Найбільш вірогідною теорією, що пояснює послідовну зміну нирок, є теорія зміни ниркових канальців. Прибічники цієї теорії вважають, що у предків хребетних була довга брунька, побудована за типом пронефроса. У їх рибообразних нащадків на спинній стороні канальців передбруньки розвинулися канальці досконалішого типа — канальці первинної бруньки, які і витіснили передниркові канальці впродовж всього тулуба, за винятком його переднього кінця. В амніот канальці вторинної бруньки, що розвивалися в задньому відділі первинної бруньки, поступово витіснили останню. Див. Виділення, Дихання органи, Шкіра .
Літ.: Холодковський Н. А., Підручник зоології і порівняльної анатомії, 3 видавництва, СП(Збори постанов) Би, 1914; Машковцев А. А., Морфологічне і філогенез відношення мезонефроса до пронефросу, «Тр. Лабораторії еволюційної морфології АН(Академія наук) СРСР», 1934, т. 2, ст 1; Керівництво по зоології, під ред. Л. А. Зенкевіча, т. 1, М. — Л., 1937; Догель Ст А., Порівняльна анатомія безхребетних, ч. 1, Л., 1938; Кравчинський Би. Д., Фізіологія нирок, [Л.], 1949; Коштоянц Х. С., Основи порівняльної фізіології, 2 видавництва, т. 1, М. — Л., 1951; Беклемішев Ст Н., Основи порівняльної анатомії безхребетних, 3 видавництва, т. 1—2, М., 1964; Курс зоології, під ред. Би. С. Матвєєва, 7 видавництво, т. 1—2. М., 1966.