Выделительная система, экскреторная система, совокупность органов, выводящих из организма животных и человека избыток воды, конечные продукты обмена, соли, а также ядовитые вещества, введённые в организм или образовавшиеся в нём. У простейших легкорастворимые экскреты (аммиак, мочевина) выводятся в окружающую среду путём диффузии (морские и паразитические формы) или с помощью сократительных вакуолей, выполняющих в основном функцию осморегуляции (у пресноводных форм). У низших водных многоклеточных (губки, кишечнополостные) и у малоактивных морских животных (иглокожие) продукты обмена, в основном аммиак, диффундируют через поверхность тела и стенки полостей, связанных с окружающей средой. В частности выделительная функция у большинства беспозвоночных свойственна кишечнику.
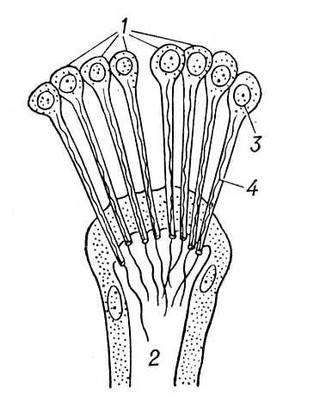
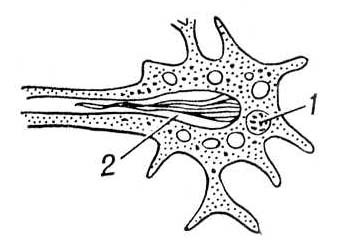
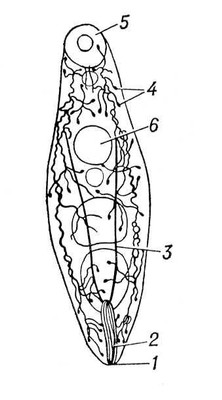
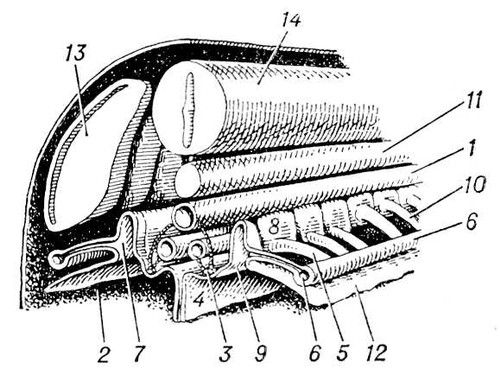
В процессе эволюции дифференцируется специальная В. с. У низших червей (плоские, гастротрихи, киноринхи, коловратки, скребни, немертины), а также у приапулид и некоторых аннелид и у личинок полихет и моллюсков выделительные органы — протонефридии — система трубочек, открывающихся наружу порами, а внутри слепо заканчивающихся особыми полыми клетками, в которых находятся пучки длинных слипшихся ресничек, непрерывно совершающих колебательные движения («мерцательное пламя») (рис. 1, 2). У кольчатых червей слепые концы протонефридиальных каналов заканчиваются группами бутылковидных клеток (соленоцитов), в узкой части которых находится по одному жгутику (рис. 3). У большинства кольчатых червей В. с. — метанефридии, т. е. метамерно расположенные парные трубчатые эктодермальные органы, открывающиеся в целомические мешки ресничной воронкой, а наружу — порой на поверхности следующего сегмента. Если в состав таких органов выделения входят и мезодермальные элементы, то говорят о нефромиксии. Выделительные органы, развивающиеся из мезодермы, — так называемые целомодукты — имеются у моллюсков: воронка открывается в околосердечную полость (остаток целома), наружное отверстие — в мантийную полость. У ракообразных выделительные органы — извитые трубки, начинающиеся слепым «целомическим» мешочком и открывающиеся наружу у основания антенн или максилл (отсюда названия «антеннальные» и «максиллярные» железы) (рис. 4). Сходны выделительные органы у мечехвостов («коксальные железы»), открывающиеся у основания 5-й пары ног. Открывающиеся наружу выделительные органы, через которые выводятся легко растворимые продукты обмена, типичны для водных беспозвоночных.
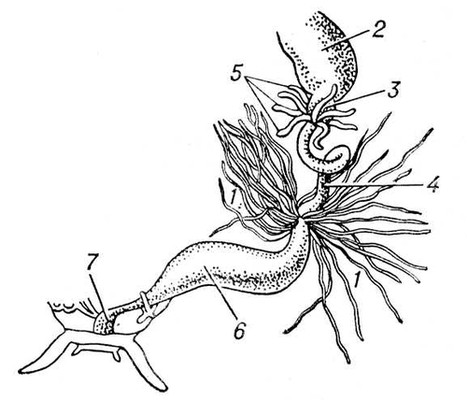
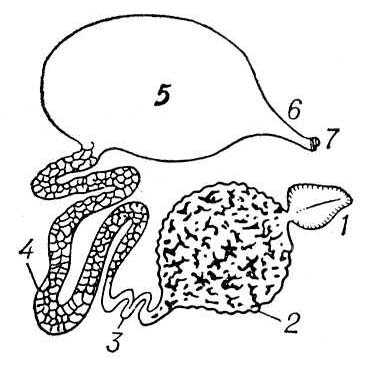
Обитание на суше, требующее экономии расхода влаги, вызывает изменение характера катаболитов — легко растворимые аммиак и мочевина заменяются трудно растворимыми гуанином (у паукообразных) или мочевой кислотой (у многоножек и насекомых). Выделительные органы типа целомодуктов из наземных беспозвоночных имеют только влаголюбивые формы — онихофоры и некоторые паукообразные (например, сенокосцы). Для наземных членистоногих характерен переход выделительной функции к стенкам кишечника (становятся иногда единственным выделительными органом, например, у ложноскорпионов) или чаще к мальпигиевым сосудам — длинным слепым выростам кишечника на границе средней и задней кишки, образующимся как выросты средней (паукообразные) или задней (насекомые, многоножки) кишки (рис. 5). Физиологическое преимущество мальпигиевых сосудов (при обитании в условиях дефицита влаги) в том, что через них легко выпадающие в осадок продукты обмена (мочевая кислота и др.) выводятся с мочевой жидкостью не наружу, а в заднюю кишку, где происходит всасывание воды; обезвоженные экскреты вместе с непереваренными остатками пищи выводятся через анальное отверстие наружу, чем достигается сохранение воды в теле животного.
Своеобразный тип выделения — отложение переведённых в труднорастворимую форму катаболитов (мочевая кислота и др.) в специальных клетках — «почках накопления» у нематод, в хлорагогенных клетках у дождевых червей, в межуточной ткани у пауков, в жировом теле у насекомых и многоножек, в печёночных выростах у мокриц и т.п., что тоже связано с экономией расхода воды при экскреции. Сходны с этими выделительными органами и экскретофоры — клетки, накапливающие экскреты и выбрасывающие их, например, в просвет кишечника (у ложноскорпионов, у восьмилучевых кораллов и др.).
Вторичный переход наземных беспозвоночных к жизни в воде, например, личинок насекомых, привёл к возвращению экскретов к характерному для водных животных типу (основной катаболит — аммиак) и к возрастанию осморегуляторной функции выделительных органов (мальпигиевы сосуды и кишечник).
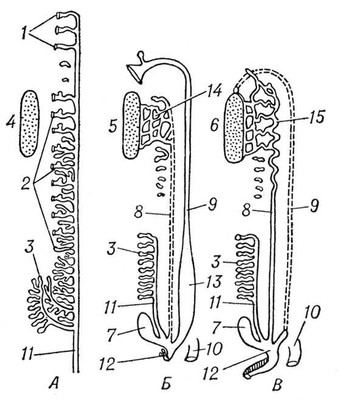
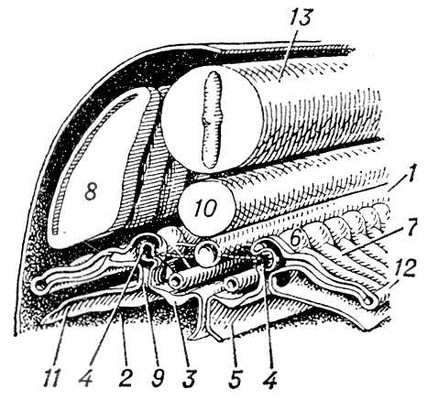
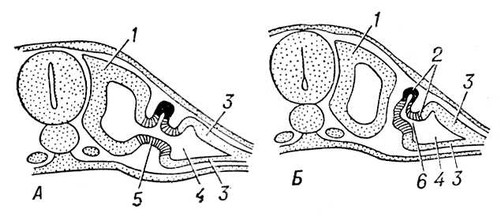
Среди хордовых у оболочников и асцидий органами выделения служат мешочки накопления (у некоторых оболочников выделительные элементы не обнаружены). У ланцетника в жаберной области имеется около 100 пар так называемых нефромиксиев, которые одним отверстием открываются в околожаберную полость, а несколькими другими отверстиями (густо усаженными соленоцитами) связаны с полостью тела. У позвоночных органами выделения служат типичные целомодукты, скопления которых образуют почки. От почек отходят мочеточники, которые впадают непосредственно в клоаку или в мочевой пузырь, открывающийся наружу мочеиспускательным каналом. В эволюционном ряду позвоночных, как и при индивидуальном развитии высших позвоночных, наблюдается последовательная смена трёх типов почек. Первыми возникают передние, или головные, почки, называемые также предпочками (см. Пронефрос). Далее развиваются первичные, средние, или туловищные, почки — вольфовы тела (см. Мезонефрос). Последними появляются вторичные, задние, или тазовые, почки (см. Метанефрос). Целомодукты предпочки и первичной почки, обычно по паре на сегмент, возникают из так называемого нефротома. Часть нефротома, открывающаяся во вторичную полость тела, образует мерцательную воронку (нефростом). Против нефростомов предпочки в полости тела обычно развивается общий для всех воронок клубок кровеносных сосудов; через него фильтруются из крови жидкие продукты выделения (водные растворы солей), которые и поступают в воронки. Предпочка, имеющая сегментарное строение и тянущаяся вдоль всего туловища, сохраняется в продолжении всей жизни из круглоротых только у бделлостомы. У миног остатки пронефроса, образующие передний отдел почки, открываются в перикардиальную полость. У других позвоночных пронефрос служит лишь личиночным органом. Он особенно хорошо развит у личинок большинства рыб и земноводных. У всех рыб и земноводных во взрослом состоянии функционируют первичные почки. Канальцы их развиваются у эмбриона сходно с канальцами предпочки, отличаясь лишь тем, что открываются в готовый пронефрический проток, который получает с этого времени название первичнопочечного канала (рис. 7 и 8). У акуловых рыб этот канал расщепляется с переднего конца на два протока: вольфов канал, служащий мочеточником первичной почки, и мюллеров канал, вступающий в связь с остатками пронефроса и выполняющий у самок функцию яйцевода. У самцов земноводных вольфов канал находится в связи с половой железой и выполняет одновременно функции и мочеточника, и семяпровода. Для первичной почки характерно наличие мальпигиевых телец. У амниот вторичная почка заменяет функционирующую в эмбриональном периоде первичную почку, которая у взрослых самок вообще дегенерирует, а у самцов несёт функцию семяпроводящего аппарата и называется придатком семенника, или эпидидимисом. Эмбрионально вторичная почка образуется из специального мочеточника, вырастающего из заднего конца вольфова канала (рис. 9). Этот мочеточник имеет на конце несколько конечных разветвлений, в которые впадают почечные канальцы, возникающие из нефрогенной ткани. Извитые канальцы вторичной почки никогда не имеют воронок и начинаются боуменовой капсулой. У млекопитающих и человека за капсулой следуют проксимальные извитые канальцы, далее прямые нисходящие и восходящие канальцы, образующие так называемую петлю Генле, и, наконец, дистальные извитые канальцы и затем соединительные трубки, впадающие в выводной проток. Расширенная часть мочеточника, откуда отходят его конечные разветвления в почку, получает назв.(название) почечной лоханки. Три типа почек связаны между собой известными переходами. Так, даже в предпочке могут быть отдельные клубочки, свойственные первичной почке. В последней довольно часто исчезают воронки, и она становится очень похожей по строению на вторичную почку.
Наиболее вероятной теорией, объясняющей последовательную смену почек, является теория смены почечных канальцев. Сторонники этой теории считают, что у предков позвоночных была длинная почка, построенная по типу пронефроса. У их рыбообразных потомков на спинной стороне канальцев предпочки развились канальцы более совершенного типа — канальцы первичной почки, которые и вытеснили предпочечные канальцы на протяжении всего туловища, за исключением его переднего конца. У амниот канальцы вторичной почки, развивавшиеся в заднем отделе первичной почки, постепенно вытеснили последнюю. См. Выделение, Дыхания органы, Кожа.
Лит.: Холодковский Н. А., Учебник зоологии и сравнительной анатомии, 3 изд., СП(Собрание постановлений)Б, 1914; Машковцев А. А., Морфологическое и филогенетическое отношение мезонефроса к пронефросу, «Тр. Лаборатории эволюционной морфологии АН(Академия наук) СССР», 1934, т. 2, в. 1; Руководство по зоологии, под ред. Л. А. Зенкевича, т. 1, М. — Л., 1937; Догель В. А., Сравнительная анатомия беспозвоночных, ч. 1, Л., 1938; Кравчинский Б. Д., Физиология почек, [Л.], 1949; Коштоянц Х. С., Основы сравнительной физиологии, 2 изд., т. 1, М. — Л., 1951; Беклемишев В. Н., Основы сравнительной анатомии беспозвоночных, 3 изд., т. 1—2, М., 1964; Курс зоологии, под ред. Б. С. Матвеева, 7 изд., т. 1—2. М., 1966.