Почки, важнейшие парные органы выделения позвоночных животных и человека, участвующие в водно-солевом гомеостазе, т. е. в поддержании постоянства концентрации осмотически активных веществ в жидкостях внутренней среды (см. Осморегуляция), постоянства объёма этих жидкостей, их ионного состава и кислотно-щелочного равновесия. Через П. выводятся из организма конечные продукты азотистого обмена, чужеродные и токсические соединения, избыток органических и неорганических веществ. П, участвуют в метаболизме углеводов и белков, в образовании биологически активных веществ, регулирующих уровень артериального давления, скорость секреции альдостеронанадпочечниками и скорость образования эритроцитов.
Сравнительная морфология. В процессе эволюции позвоночных животных происходила последовательная смена трёх типов П.: предпочки, или головной П. (пронефрос), первичной, или туловищной, П. (мезонефрос), вторичной, или тазовой, П. (метанефрос). Эти типы П. сменяют друг друга и в течение индивидуального развития высших позвоночных. Все типы П. развиваются из ножки сомита — нефротома. Первичная П. — орган выделения, функционирующий в послезародышевый период у круглоротых, рыб, земноводных, молодых ящериц и у некоторых млекопитающих (клоачные, сумчатые) до наступления у них половой зрелости. У всех остальных позвоночных и у человека первичная П. сменяется вторичной ещё у зародышей. Основная морфо-функциональная единица П. — нефрон. В процессе эволюции П. у позвоночных происходило развитие структур нефрона и их видоизменение, связанное с приспособлением животных к различным средам. У круглоротых (миноги) все нефроны начинаются из общей полости гломуса, в которую происходит ультрафильтрация из кровеносных капилляров. У рыб и остальных позвоночных имеется уже мальпигиево тельце, которым начинается каждый нефрон. Только у некоторых видов морских костистых рыб нефроны не имеют клубочков. В П. всех позвоночных нефрон имеет проксимальный сегмент, а у подавляющего большинства животных (кроме нескольких видов морских костистых рыб) и дистальный. У птиц и млекопитающих развивается новая морфология, структура — петля Генле, канальцы которой служат основным элементом мозгового вещества П.
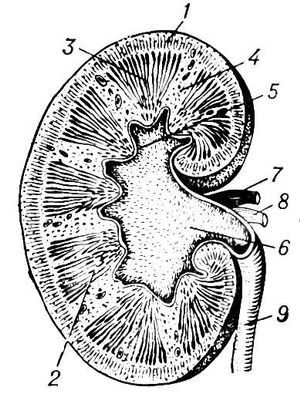
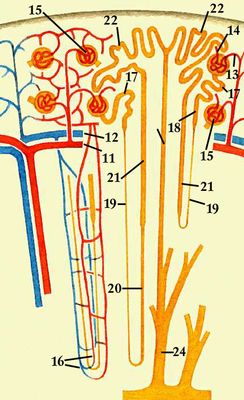
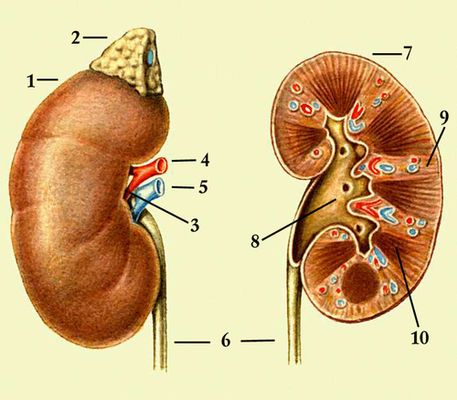
У человека П. — парные бобовидные органы, расположенные на задней брюшной стенке по обеим сторонам позвоночника обычно на уровне 12-го грудного — 3-го поясничного позвонков (рис. 1). Известны аномалии развития, когда имеется 1 или 3 П. У взрослого человека каждая П. весит 120—200 г, её длина 10—12 см, ширина 5—6 см, толщина 3—4 см. Передняя поверхность П. покрыта брюшиной, но сама П. находится вне брюшинной полости. П. окружены фасцией, под которой находится жировая капсула; непосредственно паренхима П. окружена фиброзной капсулой. П. имеет гладкий выпуклый наружный край и вогнутый внутренний край, в центре его находятся ворота П., через которые открывается доступ в почечную пазуху с почечной лоханкой, продолжающейся в мочеточник. В этом же месте в П. входят артерия и нервы; выходят вена и лимфатические сосуды. Отличительной особенность П. млекопитающих — ясно выраженное деление на 2 зоны — внешнюю (корковую) красно-коричневого цвета и внутреннюю (мозговую), имеющую лилово-красный цвет (рис. 2). Мозговое вещество П. образует 8—18 пирамид; над пирамидами и между ними лежат слои коркового вещества — почечные (бертиниевы) столбы. Каждая пирамида имеет широкое основание, примыкающее к корковому веществу, и закруглённую и более узкую верхушку — почечный сосочек, обращенный в малую почечную чашечку. Последние открываются в большие почечные чашечки, из них моча поступает в почечную лоханку и далее в мочеточник (см. Мочевыделение). В обеих П. человека около 2 млн. нефронов. Каждый нефрон состоит из частей, имеющих характерное строение и выполняющих неодинаковую функцию. Начальная часть нефрона (боуменова капсула) в виде чаши окружает сосудистый клубочек Шумлянского, образуя вместе мальпигиево, или почечное, тельце. Боуменова капсула продолжается в проксимальный извитой мочевой каналец, переходящий в прямую часть проксимального канальца. За ним следует тонкий нисходящий участок петли Генле, спускающийся в мозговое вещество П., где он, изгибаясь на 180°, переходит в тонкий восходящий, а затем толстый восходящий каналец петли Генле, возвращающийся к клубочку. Восходящая часть петли переходит в дистальный (вставочный) отдел нефрона; он соединяется связующим отделом с расположенными в коре П. собирательными трубками. Они проходят корковое и мозговое вещество П. и, сливаясь вместе, образуют в сосочке беллиниевы протоки, открывающиеся в почечную лоханку. В П. млекопитающих и человека имеется несколько типов нефронов, различающихся по месту расположения клубочков в коре П. и по функции канальцев: субкортикальные, интеокортикальные и юкстамедуллярные. Клубочки субкортикальных нефронов находятся в поверхностной зоне коры П., юкстамедуллярные — у границы коркового и мозгового вещества П. Юкстамедуллярные нефроны имеют длинную петлю Генле, глубоко спускающуюся в почечный сосочек и обеспечивающую высокий уровень осмотического концентрирования мочи. Для П. характерно строгое зональное распределение различных типов канальцев. В коре П. находятся все клубочки, проксимальные и дистальные извитые канальцы, корковые отделы собирательных трубок. В мозговом веществе располагаются петли Генле и собирательные трубки. От расположения отдельных элементов нефрона зависит эффективность осморегулирующей функции П.
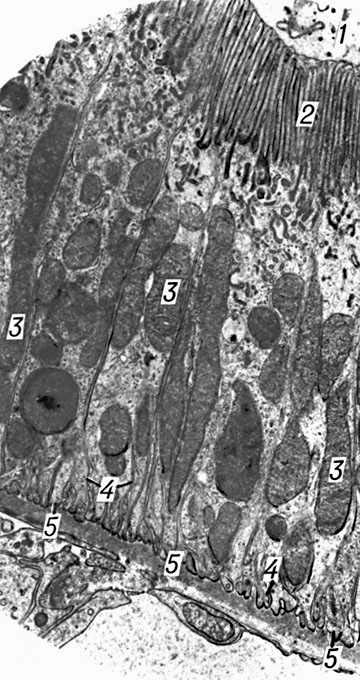
Клетки каждого отдела канальцев отличаются по строению. Для кубического эпителия проксимального извитого канальца характерны многочисленные микроворсинки (щёточная каёмка) на поверхности, обращенной в просвет нефрона. На базальной поверхности клеточная оболочка образует узкие складки, между которыми расположены многочисленные митохондрии (рис. 3, а). В клетках прямого участка проксимального канальца менее выражены щёточная каёмка и складчатость базальной мембраны, мало митохондрии. Тонкий отдел петли Генле меньшего диаметра, выстлан плоскими клетками с малочисленными митохондриями. Характерная особенность эпителия дистального сегмента нефрона (толстый восходящий отдел петли Генле и дистальный извитой каналец со связующим отделом) — малое число микроворсинок на поверхности канальца, обращенной в просвет нефрона, ярко выраженная складчатость базальной плазматической мембраны и многочисленные крупные митохондрии с большим числом крист (рис. 3, б). В начальных отделах собирательных трубок чередуются светлые и тёмные клетки (в последних больше митохондрии). Беллиниевы трубки образованы высокими клетками с немногочисленными митохондриями.
Кровь в П. поступает из брюшной аорты по почечной артерии, распадающейся в ткани П. на междолевые, дуговые, междольковые артерии, от которых берут начало афферентные (приносящие) артериолы клубочков. В них артериола распадается на капилляры, затем они вновь соединяются, образуя эфферентную (выносящую) артериолу. Афферентная артериола почти в 2 раза толще эфферентной, что способствует клубочковой фильтрации. Эфферентная артериола вновь распадается на капилляры, оплетающие канальцы того же самого нефрона. Венозная кровь поступает в междольковые, дуговые и междолевые вены; они образуют почечную вену, впадающую в нижнюю полую вену. Кровоснабжение мозгового вещества П. обеспечивается прямыми артериолами. П. иннервируют симпатические нейроны трёх нижних грудных и двух верхних поясничных сегментов спинного мозга; парасимпатические волокна идут к П. от блуждающего нерва. Чувствительная иннервация П, в составе чревных нервов достигает нижних грудных и верхних поясничных узлов.
Сравнительная физиология. Основные функции П. (экскреторная, осмо- и ионорегулирующая и др.) обеспечиваются процессами, лежащими в основе мочеобразования: ультрафильтрацией жидкости и растворённых веществ из крови в клубочках, обратным всасыванием частиц этих веществ в кровь и секрецией некоторых веществ из крови в просвет канальца. В процессе эволюции П. фильтрационно-реабсорбционный механизм мочеобразования всё более преобладает над секреторным. Регуляция выделения большинства ионов у наземных позвоночных основана на изменении уровня реабсорбции ионов. Характерная особенность эволюции П. — увеличение объёма клубочковой фильтрации, которая у млекопитающих в 10—100 раз выше, чем у рыб и земноводных; резко возрастает интенсивность реабсорбции веществ клетками канальцев, т.к. отношение массы П. к массе тела почти одинаково у этих животных. Повышается функция П. по поддержанию стабильности состава веществ, растворённых в сыворотке крови. У некоторых низших позвоночных более развит аппарат секреции, что позволяет выделять ряд веществ с такой же скоростью, как у млекопитающих. У морских рыб П. секретирует магний, сульфаты, фосфаты и т.д. В П. всех позвоночных, кроме млекопитающих и круглоротых, имеется почечно-портальная система кровообращения, которая приносит непосредственно к канальцам, минуя клубочки, венозную кровь от нижних конечностей; клетки канальцев извлекают из неё многие вещества и выделяют их путём секреции в просвет нефрона. Развитие осморегулирующей функции П. тесно связано с типом азотистого обмена. У земноводных и млекопитающих конечный продукт азотистого обмена — мочевина, осмотически высокоактивное вещество, для выведения которого необходимо значительное количество воды (земноводные) или способность осмотически концентрировать мочу (млекопитающие). У пресмыкающихся и птиц конечный продукт обмена — мочевая кислота, секретируемая клетками почечных канальцев; при реабсорбции воды образуются пересыщенные растворы; в клоаке мочевая кислота, плохо растворимая в воде, выпадает в осадок — это позволяет организму сберегать воду. Для выделения мочевины, образовавшейся из 1 г белка, необходимо 20 мл воды (при равенстве осмотических концентраций мочи и плазмы крови), а для экскреции соответствующего количества мочевой кислоты — 0,5 мл.
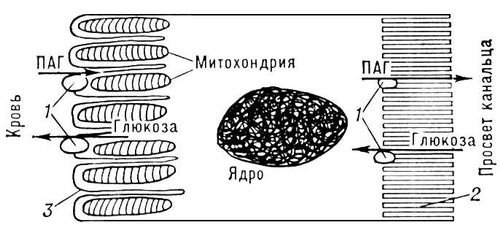
У человека в условиях покоя около 1/4 части крови, выбрасываемой в аорту левым желудочком сердца, поступает в почечные артерии. Кровоток в П. мужчин составляет 1300 мл/мин, у женщин несколько меньше. При этом в клубочках из полости капилляров в просвет боуменовой капсулы происходит ультрафильтрация плазмы крови, обеспечивающая образование т. н. первичной мочи, в которой практически нет белка. В просвет канальцев поступает около 120 мл жидкости в 1 мин. Однако в обычных условиях около 119 мл фильтрата поступает обратно в кровь и лишь 1 мл в виде конечной мочи выводится из организма. Процесс ультрафильтрации жидкости обусловлен тем, что гидростатическое давление крови в капиллярах клубочка выше суммы коллоидно-осмотическое давления белков плазмы крови и внутрипочечного тканевого давления. Размер частиц, фильтруемых из крови, определяется величиной пор в фильтрующей мембране, что, по-видимому, зависит от диаметра пор центрального слоя базальной мембраны клубочка. В большинстве случаев радиус пор меньше 28 , поэтому электролиты, низкомолекулярные неэлектролиты и вода свободно проникают в просвет нефрона, белки же практически не проходят в ультрафильтрат, функциональное значение отдельных почечных канальцев в процессе мочеобразования неодинаково. Клетки проксимального сегмента нефрона всасывают (реабсорбируют) попавшие в фильтрат глюкозу (рис. 4), аминокислоты, витамины, большую часть электролитов. Стенка этого канальца всегда проницаема для воды; объём жидкости к концу проксимального канальца уменьшается на 2/3, но осмотическая концентрация жидкости остаётся той же, что и плазмы крови. Клетки проксимального канальца способны к секреции, т. е. выделению некоторых органических кислот (пенициллин, кардиотраст, парааминогиппуровая кислота, флуоресцеин и др.) и органических оснований (холин, гуанидин и др.) из околоканальцевой жидкости в просвет канальца. Клетки дистального сегмента нефрона и собирательных трубок участвуют в реабсорбции электролитов против значительного электрохимического градиента; некоторые вещества (калий, аммиак, ионы водорода) могут секретироваться в просвет нефрона. Проницаемость стенок дистального извитого канальца и собирательных трубок для воды увеличивается под влиянием антидиуретического гормона — вазопрессина, вследствие чего происходит всасывание воды по осмотическому градиенту.
Осморегулирующая функция П. обеспечивает постоянство концентрации осмотически активных веществ в крови при различном водном режиме. При избыточном поступлении воды в организм выделяется гипотоническая моча, в условиях дефицита воды образуется осмотически концентрированная моча. Механизм осмотического разведения и концентрирования мочи был открыт в 50—60-х гг. 20 в. В П. млекопитающих канальцы и сосуды мозгового вещества образуют противоточно-поворотную множительную систему. В мозговом веществе П. параллельно друг другу проходят нисходящие и восходящие отделы петель Генле, прямые сосуды, собирательные трубки. В результате активного транспорта натрия клетками восходящего отдела петли Генле соли натрия накапливаются в мозговом веществе П. и вместе с мочевиной удерживаются в этой зоне П. При движении крови вниз, в глубь мозгового вещества, мочевина и соли натрия поступают в сосуды, а при обратном движении, к корковому веществу, выходят из них, удерживаясь в ткани (принцип противотока). При действии вазопрессина высокая осмотическая концентрация характерна для всех жидкостей (кровь, межклеточная и канальцевая жидкость) на каждом уровне мозгового вещества П., исключая содержимое восходящих отделов петель Генле. Стенки этих канальцев относительно водонепроницаемы, а клетки активно реабсорбируют соли натрия в окружающую межуточную ткань, вследствие чего осмотическая концентрация уменьшается. При отсутствии вазопрессина стенка собирательных трубок водонепроницаема; при действии этого гормона она становится водопроницаемой и вода всасывается из просвета по осмотическому градиенту в окружающую ткань. В П. человека моча может быть в 4—5 раз осмотически концентрированнее крови. У некоторых обитающих в пустынях грызунов, имеющих особенно развитое внутреннее мозговое вещество П., моча может в 18 раз превосходить по осмотическому давлению кровь.
Изучены молекулярные механизмы реабсорбции и секреции веществ клетками почечных канальцев. При реабсорбции натрий пассивно поступает по электрохимическому градиенту внутрь клетки, движется по ней к области базальной плазматической мембраны и с помощью находящихся в ней «натриевых насосов» (Na/K ионообменный насос, электрогенный Na насос и др.) выбрасывается во внеклеточную жидкость. Каждый из этих насосов угнетается специфическими ингибиторами. Применение в клинике мочегонных средств, используемых, в частности, при лечении отёков, основано на том, что они влияют на различные элементы системы реабсорбции Na. К, в отличие от Na, клетка нефрона может не только реабсорбировать, но и секретировать. При секреции К из межклеточной жидкости поступает в клетку через базальную плазматическую мембрану за счёт работы Na/K насоса, а выделяется он в просвет нефрона через апикальную клеточную мембрану пассивно. Это обусловлено увеличением калиевой проницаемости мембран и высокой внутриклеточной концентрацией К. Реабсорбция различных веществ регулируется нервными и гормональными факторами. Всасывание воды возрастает под влиянием вазопрессина, реабсорбция Na увеличивается альдостероном и уменьшается натрийуретическим фактором, всасывание Ca и фосфатов изменяется под влиянием паратиреоидного гормона и тирокальцитонина (см. Щитовидная железа) и др. Молекулярные механизмы регуляции переноса различных веществ клеткой нефрона неодинаковы. Так, ряд гормонов (например, вазопрессин) стимулирует внутриклеточное образование из АТФ циклические формы АМФ, которая воспроизводит эффект гормона. Другие же гормоны (например, альдостерон) воздействуют на генетический аппарат клетки, вследствие чего в рибосомах усиливается синтез белков, обеспечивающих изменение переноса веществ через клетку канальца.
Важное значение имеет П. как инкреторный (внутрисекреторный) орган. В клетках её юкстагломерулярного аппарата, расположенного в области сосудистого полюса клубочка между приносящей и выносящей артериолами, происходит образование ренина, а возможно, и эритропоэтина. Секреция ренина возрастает при уменьшении почечного артериального давления и снижении содержания Na в организме (см. Ренин-ангиотензинная система). В П. вырабатывается как эритропоэтин, так и, по-видимому, вещество, угнетающее образование эритроцитов; эти вещества участвуют в регуляции эритроцитарного состава крови. Установлено, что в П. синтезируются простагландины, вещества, меняющие чувствительность почечной клетки к некоторым гормонам (например, вазопрессину) и снижающие кровяное давление. См. также Водно-солевой обмен, Выделительная система, Мочеполовая система.
Лит.: Иванов Г. Ф., Основы нормальной анатомии человека, т. 1, М., 1949; Гинецинский А. Г., Физиологические механизмы водно-солевого равновесия, М. — Л., 1963; Основы нефрологии, под ред. Е. М. Тареева, т. 1—2, М., 1972; Физиология почки, под ред. Ю. В. Наточина, Л., 1972; Handbuch (der mikroskopischen Anatomic des Menschen, Bd 7, Tl 1—2, В., 1930; Smith H. W., The kidney. Structure and function in health and disease, N. Y., 1951; Wesson L. G., Physiology of the human kidney, N. Y. — L., 1969; The kidney, morphology, biochemistry, physiology, ed. C. Rouiller, A. F. Muller, v. 1—4, N. Y. — L., 1969—71.
Ю. В. Наточин.
Болезни почек могут проявляться изменениями мочи, отёками, гипертонией и др. симптомами. Различают врождённые и приобретённые заболевания П. К врождённым относятся аномалии числа (единственная, добавочная, раздвоенная П.), положения (опущение, высокое расположение П., одностороннее положение П.), формы (подково-, галетообразная П., S-образная, L-образная) и структуры (аплазия, гипоплазия, поликистоз П. и др.) П.; различные аномалии почечной лоханки и мочеточников (числа, формы, калибра и др.); генетически обусловленные пороки анатомического развития и строения почечной паренхимы, нередко сопровождающиеся врождёнными дефектами др. органов (например, семейная нефропатия с глухотой); тубулопатии — заболевания почечных канальцев, в основном связанные с нарушениями обмена веществ (аминокислот, ферментов — например семейная цистинурия, витамин-D-резистентный рахит и др.).
Среди приобретённых болезней П. наиболее часты аутоиммунные заболевания, например гломерулонефрит (см. Нефрит)или нефропатия беременных, и инфекционные — пиелонефрит, туберкулёз и др. Нередки поражения П. токсического характера: при отравлениях тяжёлыми металлами и их соединениями (например, ртутью, висмутом и др.), пестицидами, органическими растворителями (тетрахлорметан, тетрахлорэтилен и др.); лекарственные нефропатии (при приёме сульфаниламидов, мочегонных и др.); радиационные, травматические повреждения и др. Опухоли П. бывают злокачественными (гипернефроидный рак, саркома) и доброкачественными (фиброма, миксома, аденома и др.), первичными и метастатическими. Часто встречается почечнокаменная болезнь. Поражения П. могут быть сопутствующими — вторичными по отношению к заболеваниям др. органов и систем (коллагеновым болезням, системным васкулитам, сахарному диабету и т.д.). Нередко сопутствующие нефропатии определяют тяжесть и исход основного заболевания.
При ряде заболеваний, как почечных, так и внепочечных (шок, отравление, острая инфекция и т.д.), могут внезапно нарушаться основная функции П.: возникает острая почечная недостаточность, характеризующаяся повышением содержания азота в крови, нарушением водно-электролитного баланса и кислотно-щелочного равновесия. Хроническая почечная недостаточность может быть исходом многих неизлеченных заболеваний П. Однотипность проявлений различных по своему происхождению и характеру заболеваний П., а также возможность различных клинических проявлений одного и того же заболевания явились основанием для выделения ряда почечных синдромов. Наряду с острой и хронической почечной недостаточностью к ним относят: нефротический синдром, почечную гипертонию (см. Симптоматические гипертонии), синдром канальцевой (тубулярной) недостаточности, ведущий к нарушению гомеостаза внутренней среды организма. Лечение болезней П. строится с учётом причины, характера (преобладание определённого синдрома) и стадии течения заболевания.