Симетрія в біології (біосиметрія). На явище С. в живій природі звернули увагу ще в Древній Греції піфагорійці (5 ст до н.е.(наша ера)) у зв'язку з розвитком ними вчення про гармонію. У 19 ст з'явилися одиничні роботи, присвячені С. рослин (французькі учені О. П. Декандоль, О. Браво), тварин (німецький — Е. Геккель), біогенних молекул (французькі — А. Вешан, Л. Пастер і ін.). У 20 ст біооб'єкти вивчали з позицій загальної теорії С. (радянські учені Ю. Ст Вульф, Ст Н. Беклемішев, Би. До. Вайнштейн, голландський фізікохимік Ф. М. Егер, англійський кристалографи на чолі з Дж. Берналом) і учення про правізне і лівизну (радянські вчені Ст І. Вернадський, Ст Ст Алпатов, Г. Ф. Гаузе і др.; німецьке вчене Ст Людвіг). Ці роботи привели до виділення в 1961 особливого напряму в ученні о С. — біосимметріки.
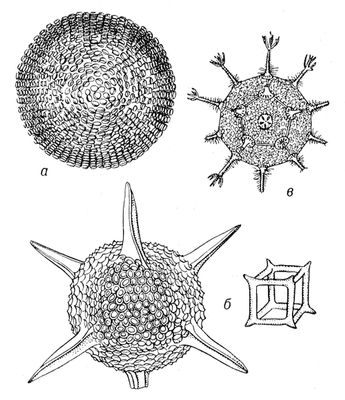
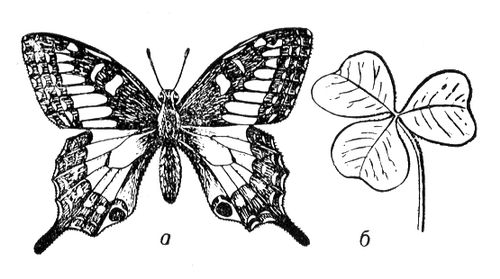
найінтенсивніше вивчалася структурна С. біооб'єктів. Дослідження С. біоструктур — молекулярних і надмолекулярних — з позицій структурною С. дозволяє заздалегідь виявити можливі для них види С., а тим самим число і вигляд можливих модифікацій, строго описувати зовнішню форму і внутрішню будову будь-яких просторових біооб'єктів. Це привело до широкого використання вистав структурною С. в зоології, ботаніці, молекулярній біології. Структурна С. виявляється перш за все у вигляді того або іншого закономірного повторення. У класичній теорії структурної С., розвиненою німецьким ученим І. Ф. Гесселем, Е. С. Федоровим і іншими, вигляд С. об'єкту може бути описаний сукупністю елементів його С., тобто таких геометричних елементів (крапок, ліній, плоскості), відносно яких впорядковані однакові частини об'єкту (див. Симетрія в математиці). Наприклад, вигляд С. квітки флокса ( мал. 1 , в) — одна вісь 5-го порядку, що проходить через центр квітки; вироблювані за допомогою її операції — 5 поворотів (на 72, 144, 216, 288 і 360°), при кожному з яких квітка збігається з самим собою. Вигляд С. фігури метелика ( мал. 2 , би) — одна плоскість, що ділить її на 2 половини, — ліву і праву; вироблювана за допомогою плоскості операція — дзеркальне віддзеркалення, що «робить» ліву половинку правою, праву — лівою, а фігуру метелика що поєднує з самою собою. Вигляд С. радіолярії Lithocubus geometricus ( мал. 3 , би), окрім осей обертання і плоскості віддзеркалення містить ще і центр С. Любая проведена через таку єдину крапку усередині радіолярії пряма по обидві сторони від неї і на рівних відстанях зустрічає однакові (відповідні) точки фігури. Операції, вироблювані за допомогою центру С., — віддзеркалення в крапці, після яких фігура радіолярії також поєднується сама з собою.
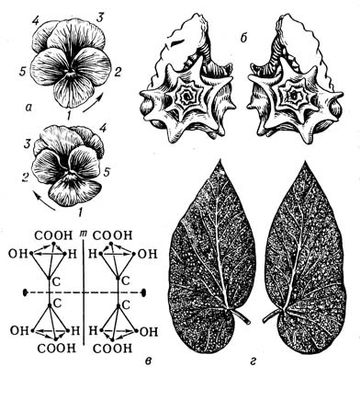
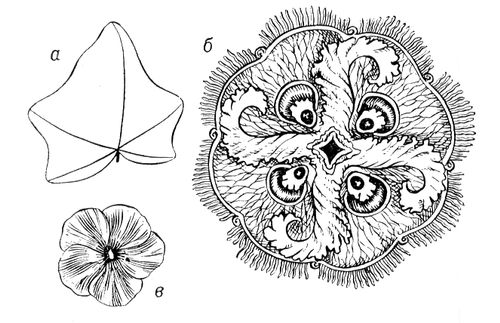
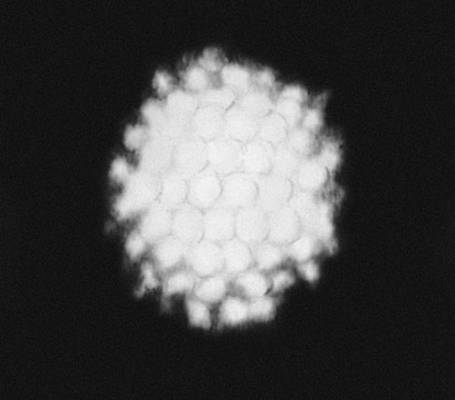
В живій природі (як і в неживій) із-за різних обмежень зазвичай зустрічається значно менше число видів С., чим можливо теоретично. Наприклад, на нижчих етапах розвитку живої природи зустрічаються представники всіх класів точкової С. — аж до організмів, що характеризуються С. правильних многогранників і кулі (див. мал. 3 ). Проте на вищих рівнях еволюції зустрічаються рослини і тварини в основному т.з. аксіальною (вигляду n ) і актиноморфною (вигляду n ( m ) З . (у обох випадках n може набувати значень від 1 до ¥). Біооб'єкти з аксіальною С. (див. мал. 1 ) характеризуються лише віссю С. порядку n . Біооб'єкти сактіноморфной С. (див. мал. 2 ) характеризуються однією віссю порядку n і пересічною по цій осі плоскістю m . У живій природі найбільш поширені С. вигляду n = 1 і 1× m = m , називається відповідно асиметрією і двосторонньою, або білатеральною, С. Асимметрія характерна для листя більшості видів рослин, двостороння С. — до певної міри для зовнішньої форми тіла людини, хребетних тварин і багатьох безхребетних. В рухливих організмів така С., мабуть, пов'язана з відмінностями їх русі вгору-вниз і вперед-назад, тоді як їх рухи направо-наліво однакові. Порушення у них білатеральною С. неминуче привело б до гальмування руху однієї із сторін і перетворенню поступальної ходи на круг. У 50—70-х рр. 20 ст інтенсивному вивченню (перш за все в СРСР) піддалися т.з. діссимметрічеськие біооб'єкти ( мал. 4 ). Останні можуть існувати принаймні в двох модифікаціях — у формі оригінала і його дзеркального віддзеркалення (антипода). При цьому одна з цих форм (неважливо яка) називається правою або D (від латів.(латинський) dextro), інша — лівою або L (від латів.(латинський) laevo). При вивченні форми і будови D- і L-біооб'ектов була розвинена теорія діссимметрізующих чинників, що доводить можливість для будь-якого D- або l-об'єкту два і більш (до безконечного числа) за модифікації (див. також мал. 5 ); одночасно в ній містилися і формули для визначення числа і вигляду останніх. Ця теорія привела до відкриття т.з. біологічною ізомерії (різних біооб'єктів одного складу; на мал. 5 змальовано 16 ізомерів аркуша липи).
При вивченні тієї, що зустрічається біооб'єктів було встановлено, що в одних випадках переважають D-, в інших l-формі, в третіх вони представлені однаково часто. Бешаном і Пастером (40-і рр. 19 ст), а в 30-х рр. 20 ст радянським ученим Г. Ф. Гаузе і іншими було показано, що клітки організмів побудовані лише або переважно з L-amінокислот, l-білків, d-дезоксирібонуклеїновіх кислот, d-цукрів, l-алкалоїдів, D- і l-терпенів і т. д. Настільки фундаментальна і характерна межа живих клітин, названа Пастером діссимметрієй протоплазми, забезпечує клітці, як було встановлено в 20 ст, активніший обмін речовин і підтримується за допомогою складних біологічних і физико-хімічних механізмів, що виникли в процесі еволюції. Сов. учений В. В. Алпатов в 1952 на 204 видах судинних рослин встановив, що 93,2% видів рослин відносяться до типа з L-, 1,5% — з d-ходом гвинтоподібних потовщень стінок судин, 5,3% видів — до типа рацемічного (число d-судін приблизно дорівнює числу l-судін).
При вивченні D- і L-біооб'ектов було встановлено, що рівноправ'я між d-і l-формамі у ряді випадків порушено із-за відмінності їх фізіологічних, біохімічних і ін. властивостей. Подібна особливість живої природи була названа діссимметрієй життю. Так, збуджуюче вплив L-amінокислот на рух плазми в рослинних клітках в десятки і сотні разів перевершує таку ж дію їх d-форм. Багато антибіотиків (Пеніцилін, Граміцидин і ін.), D-amінокислоти, що містять, володіють більшою бактерицидністю, чим їх форми з L-amінокислотамі. Частіше гвинтоподібні l-коренеплоді цукрової, що зустрічаються буряка на 8—44% (залежно від сорту) важче і містять на 0,5—1% більше цукру, чим d-коренеплоді.
Вивчення спадкоємства ознак в D- і l-форм показало, що їх правізна або лівизна може бути спадковою, неспадковою або має характер тривалою модифікації . Це означає, що принаймні у ряді випадків правізну-лівізну організмів і їх частин можна змінити дією мутагенних або немутагенних хімічних сполук. Зокрема, d-штамі (по морфології колоній) мікроорганізму Bacillus mycoides при вирощуванні їх на агарі з d-сахарозою, L-днгитоніном, d-вінною кислотою можна перетворити на l-штамі, а l-штамі можна перетворити на d-штамі, вирощуючи їх на агарі з l-вінною кислотою і d-амінокислотамі. У природі взаємоперетворення D- і l-форм можуть відбуватися і без втручання людини. При цьому зміна видів С. в еволюції відбувалася не лише в діссимметрічеських організмів. В результаті виникли багаточисельні еволюційні ряди С., специфічні для тих або інших гілок древа життя.
Структурна С. біосистем вивчається також з точки зору загальніших типів С. — кольоровий С., С. подібності, антисиметрії і ін.
Розробка учення о С. біооб'єктів дозволить поглибити уявлення як про їх властивості і функції, так і про походження і суть життя .
Літ.: Гаузе Р. Ф., Асиметрія протоплазми, М. — Л., 1940; Вайнштейн Би. До., Дифракція рентгенових променів на ланцюгових молекулах, М., 1963; Беклемішев Ст Н., Основи порівняльної анатомії безхребетних, 3 видавництва, т. 1—2, М., 1964; Урманцев Ю. А., Симетрія природи і природа симетрії, М., 1974; Ludwig W., Das Rechts-links-problem im Tierreich und beim Menschen..., B. — Hdib. — N. Y., 1970; Bentley R., Molecular asymmetry in biology, v. 1—2, N. Y., 1969—70.