Радіочутливість, чутливість біологічних об'єктів до дії іонізуючих випромінювань . Опромінення викликає в клітках і організмах різні зміни> (див. Біологічна дія іонізуючих випромінювань ), міра прояву яких не завжди корелює між собою. Тому при оцінці Р. поважно враховувати, який критерій використовується для її характеристики. Зазвичай таким критерієм служить летальна дія випромінювань — інактівация або загибель кліток і загибель багатоклітинних організмів. Летальна дія випромінюванні також може виявлятися в різних формах: в разі кліток — загибель їх в інтерфазі після одного або декількох ділень (див. Мітоз ), в разі багатоклітинних організмів — загибель в різні терміни після опромінення.
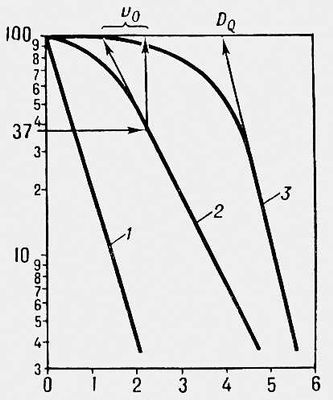
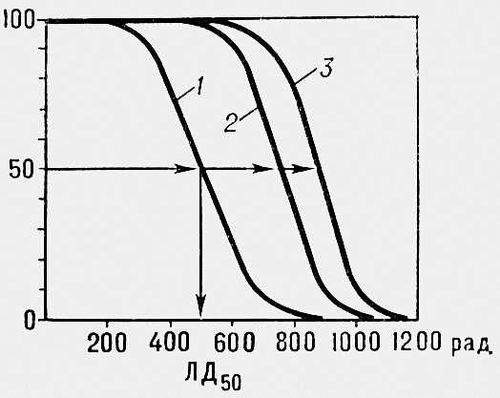
Щоб оцінити Р., біологічні об'єкти опромінюють різними дозами, визначають відсоток тих, що вижили і будують криві виживання. Для кліток такі криві змальовують зазвичай в напівлогарифмічному масштабі ( мал. 1 ), для багатоклітинних організмів — в лінійному ( мал. 2 ). Користуючись кривими виживання, знаходять ЛД 50 — дозу, після якої виживає 50% особин, а також значення D Q і D 0 , що відображають величину «плеча» і нахил прямолінійної складової таких кривих (значення D 0 дорівнює дозі, що зменшує виживаність в е 2,7 разу на прямолінійній складовій кривої виживання). У експериментах з ссавцях ЛД 50 визначають зазвичай для різних термінів після опромінення — 3, 5, 15, 30 і т.д. доби. Значень ЛД, що набувають, 50 / 5 , ЛД 50 / 30 і т.п. відображають Р. тих систем організму, переважна поразка яких відповідально за його загибель протягом того або іншого відрізання часу. Так, загибель мишей і щурів протягом перших 3—5 сут після опромінення пов'язана з пошкодженням кишкового тракту, а в інтервалі між 5 і 30 сут — з пошкодженням системи кровотворення. Мірою Р. зазвичай служать ЛД 50 або D 0 .
Р. кліток може розрізнятися в сотні і тисячі разів: ЛД 50 для кліток ссавців — 200—350 рад , для бактерій і дріжджів — 10—45 тис. рад , для інфузорій і амеб — 300—500 тис. рад. Р. обумовлюється первинною поражаємостью життєво важливих структур кліток, їх здатністю до відновлення (репарації ) і умовами культивування. У загальному випадку Р. кліток зростає із збільшенням вмісту ДНК(дезоксирибонуклеїнова кислота), числа і розмірів хромосом і зменшується із збільшенням числа хромосомних наборів (плодоїдності ). В той же час на Р. кліток впливають їх хімічний склад (наприклад, вміст ендогенних тіолов) фізіологічний стан (фаза клітинного циклу, фаза диференціювання), умови під час опромінення (можуть надавати радіозахисну або радіосенсибілізуючу дію) і умови в радіаційний для поста період (можуть сприяти або перешкоджати здійсненню репарації і прояву первинних пошкоджень). Клітки з порушеною системою репарації відрізняються підвищеному Р. Мутації в окремих генах можуть в десятки разів змінювати Р. кліток, впливаючи на різні сторони метаболізму. Т. о., Р. кліток залежить від багатьох чинників, питома вага яких в різних об'єктів різна. Р. багатоклітинних рослин і тварин також широко варіює. Так, для насіння гороху і кукурудзи ЛД 50 рівна 5—20 тис. рад , для насіння конюшини і редису — 100—250 тис. рад (для проростков цих же рослин ЛД 50 складає 250—700 рад ); для дорослих комах ЛД 50 — 30—50 тис. рад , а для ссавців — від 350—700 до 1000—1200 рад. Р. рослин і тварин обумовлюється головним чином Р. їх кліток (в разі ссавців — Р. стволових клітин їх кровотворних органів і шлунково-кишкового тракту) і чинниками, що впливають на успішність регенерації пошкоджених опроміненням органів і тканин за рахунок розмноження кліток, що вижили. На прояв Р. впливають умови вміст після опромінення, сприяючий або перешкоджаючий одужанню від променевій хворобі . Окрім біологічних особливостей і умов середовища, Р. кліток і організмів залежить від фізичних властивостей випромінювань, потужності дози і особливостей фракціонування опромінення. Розроблені способи радіосенсибілізації, тобто штучного збільшення Р. біологічних об'єктів. Вивчення різних аспектів Р. важливе для розробки ефективних методів лікування променевих пошкоджень, радіотерапії ракових пухлин, а також у випадках вживання випромінювань для радіостимуляції рослин і в штучному мутагенезі .
Літ.: Основи радіаційної біології, М., 1964; Тімофєєв-Ресовський Н. Ст, Іванов Ст І., Корогодін Ст І., Вживання принципу попадання в радіобіології, М., 1968; Кузин А. М., структурно-метаболічна гіпотеза в радіобіології, М., 1970; Акоєв І. Р., Максимов Р. До., Малишев Ст М., Променеве ураження ссавців і статистичне моделювання, М., 1972; М'ясник М. Н., Генетичний контроль радіочутливості бактерій, М., 1974.