Мохи, наземні або рідше прісноводі автотрофні рослини, що об'єднуються у відділ мохоподібних, що включає найбільш примітивні вищі рослини. М. підрозділяють на 3 класи: антоцеротовиє, печінкові і листя, або лістостебельниє. Всі вони порівняно просто організовані, багатоклітинні, переважно багатолітні (рідше — однорічні) рослини заввишки від 1 до 50 см з частковим відособленням асиміляційної, водопровідної і механічної тканин; володіють єдиним циклом розвитку, що розрізняється в межах окремих груп деякими особливостями. Цикл розвитку М. супроводиться відособленням диплоїдного спорофіту (видозміненого спорангія) на гаплоїдном гаметофіте . В результаті статеве і безстатеве покоління розвиваються спільно на одній рослині, тобто чергування поколінь у М. носить декілька умовний характер. Із спори у М. зростає гілляста, багатоклітинна нитчаста або пластинчаста протонема, здібна до фотосинтезу. Унаслідок розвитку на протонеме багаточисельних нирок для М. характерні групові форми зростання (дерновінки, куртини, подушки), що допомагає переносити несприятливі умови і сприяє вегетативному розмноженню. В одних М. з нирок зростають пластинчасті слоєвіща, в інших — радіально або двосторонньо-симетричні втечі. Слоєвіща і втечі можуть бути обох статей або одностатево; відповідно М. бувають обох статей або різностатево, однодомними і дводомними. Органи статевого розмноження — антерідії і архегонії — частіше маються в своєму розпорядженні групами серед багатоклітинних стерильних ниток — парафіз і оточені особливими листоподібними виростамі. У антерідіях (довгастих мішечках на ніжці з одношаровою оболонкою) утворюються двожгутикові сперматозоїди, для пересування яких до яйцеклітини необхідна вода. У нижній розширеній частині архегонія поміщається яйцеклітина. Запліднення і подальший розвиток зіготи відбувається в архегонії. Із зіготи за декілька місяців розвивається спорогон — орган безстатевого розмноження (спороношенія), що в значній мірі загубив самостійність. Зелене стеблеобразноє тіло молодого спорогона потім стає жовтим, коричневим або темно-червоним і диференціюється на верхню спороносну частину — коробочку і ніжнюю — ніжку із стопою, яка вростає в тканину материнської рослини. При утворенні спор з археспорія (спорогенній тканині) відбувається мейоз . Зігота і спорогон у М. діплоїдни і складають безстатеве покоління — спорофіт. Протонема, слоєвіщниє і оліственниє гаметофори гаплоїдни і відносяться до статевого покоління — гаметофіту; в основному на них лягають функції автотрофного живлення. Завдяки цьому і здібності гаметофіта до вегетативного розмноження цикл розвитку М. тривалий час може відбуватися без утворення спорофіту (в деяких видів спорогони невідомі). Гаметофіти М. морфологічно всілякі одношарові листоподібні вирости стебла (філлоїди) відрізняються великим варіюванням ознак. Спорогони ж в багатьох групах виявляють значну схожість. На стеблах оліственних М. утворюються також хлорофілоносні лісто- або ниткоподібні вирости — парафіллії, булавоподібні волоски, вторинна протонема і ниткоподібні ризоїди, які, як повстю, можуть обволікати наземні або підземні частини рослин. Ризоїдами слоєвіща і стебла прикріпляються до субстрата або щільно з'єднуються між собою. В М. встановлені апоспорія і апогамія, гібридизація і поліплоїдія, що дає підстави для визнання їх участі у відообразовательних процесах. Поряд з цим у М. поширено формування клонів, що сприяє збереженню місцевих адаптивних змін.
Антоцеротовиє М. (Anthocerotae) — своєрідна група з 2 сімейств з 6 пологами, об'єднуючими понад 300 видів, поширених переважно в тропіках, в СРСР 4—5 видів. Слоєвіще у них зазвичай лопатеве або розетковідноє із зануреними в нього антерідіямі і архегоніямі. Спорогон, як правило, витягнутий, стручковидний, здатний подовжуватися завдяки мерістеме, розташованою в його нижній частині, з колонкою в середині і з устьіцамі в зовнішніх стінках; в підстави оточений обгорткою і розкривається 2 стулками зверху вниз. Окрім спор, утворюються одно- або багатоклітинні безплідні короткі нитки — елатери, сприяючі поширенню спор. У клітках слоєвіща і спорогонов міститься 1 або декілька хроматофорів з піреноїдамі .
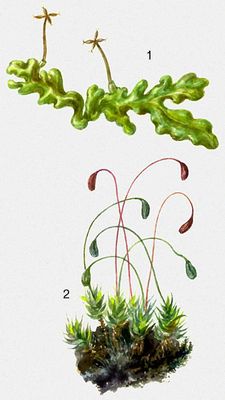
Печінкові М., або печіночники (Hepaticae), підрозділяються на 2 підкласи: маршанциевиє і юнгерманнієвиє. На відміну від інших М., у печіночників протонема слабо розвинена, недовговічна і на ній зазвичай розвивається одне слоєвіщноє або лістостебельноє рослина дорзо-вентральної структури; у клітках гаметофіта містяться одне або декілька масляних тілець постійної форми; ризоїди одноклітинні. Спорогон частіше без хлоропластів, позбавлений колонки, і на його стінках відсутні устьіца. В деяких маршанциевих виявляється схожість з антоцеротовимі, тому антоцеротових розглядали як підклас печінкових печіночників М. Гаметофіт відрізняється великою різноманітністю, спорогони однотипні. Маршанциевиє печіночники (Marchantiidae) — слоєвіщниє форми, зазвичай у вигляді маленьких розеток, що дихотомічно гілкуються, або великих стрічкоподібно слоєвіщ, що стелються, прикріпляються до грунту ризоїдами; чоловічі і жіночі гаметангиі занурені в тканину слоєвіща або підведені на особливих підставках. Переважно надгрунтові рослини, рідко скельні або водні, поширені по всій земній кулі, особливо багато представлені в тропіках. Відомо 16 сімейств з 35 пологами, об'єднуючими близько 420 видів, включаючи викопну Naiadita з мезозою. Найбільш примітивні М. з порядку сферокарпових з просто влаштованим слоєвіщем, інколи з масляними тельцями в особливих клітках, з одношаровою стінкою спорогона і з живильними клітками замість елатер. Для М. порядку маршанциевих характерна складна будова слоєвіща з розділенням на основну і асиміляційну тканину, з масляними тельцями в особливих клітках і з ськлеренхимнимі волокнами (в деяких видів). Слоєвіща часто з повітряними камерами і повітряними устьіцамі, з нижнього боку з черевними лусочками і одноклітинними гладкими і язичковими ризоїдами. Спорогон зазвичай складнішої будови. Серед них річчиевиє розглядаються як повторно спрощені, з гаметангиямі і спорогоном, що розвивається усередині дрібних, частіше розетковідних слоєвіщ. Юнгерманнієвиє печіночники (Jungermanniidae) включають групу слоєвіщних і оліственних рослин, у вегетативних клітках яких, як правило, по декілька масляних тілець; ризоїди гладкі, одноклітинні. Епіфітниє, надгрунтові і наскальні рослини особливо багато представлені в субтропіках і тропіках. 45 сімейств об'єднують понад 240 пологів що включають близько 9 тис. видів. У представників порядку метцгерієвих слоєвіще одно- або багатошарове, недиференційоване (в деяких з черевними або спинними, інколи листоподібними лусочками і стеблами з листоподібними лопатями); коробочка двух- або багатошарова. Рослини порядку гапломітрієвих прямостоячі, з трирядним листорозміщенням і коробочками, що мають одношарові бічні стінки. Найбільш багаточисельні і всілякі М. порядку юнгерманнієвих, для яких характерні 2 ряди цілісного або лопатевого бічного листя без жилки і дрібніше (інколи відсутні) черевне листя — так звані амфігастрії; стінки коробочки багатошарові.
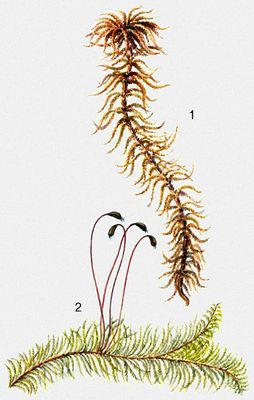
Листяні М., або лістостебельниє М. (Musci), підрозділяються на 3 підкласи: сфагнові, андрєєвиє і брієвиє. Рослини радіально-, рідко двосторонньо-симетричні, спіральнооліственниє; листя цілісне, зубчасте або по краю війчаті, одношарові, рідше дву- або багатошарові, з жилкою або без жилки; з багатоклітинними, гіллястими ризоїдами. Спорогони верхівкові або бічні, різноманітні за формою, будові і забарвленню. Близько 15 тис. видів (по ін. даним, до 25 тис. видів), що об'єднуються в 660 пологів. Сфагнові, або торф'яні, М. мають крупні, прямостоячі стебла з пучковідно розташованими гілками, на верхівці зібраними в голівку; білувато-зелені, жовті, бурі або червонуваті. Листя одношарове, гілкове і стеблове, без жилки, з хлорофілоносних і водоносних кліток, що чергуються. Спорогон складається з кулевидної коробочки з кришечкою і недорозвиненими устьіцамі, без ніжки, із стопою, врослою в безлисту подовжену архегоніальную гілочку стебла. Споровий мішок сводообразно покриває верхню частину колонки. Поширені по всій земній кулі: переважають на болотах, в тундрі, у вологих лісах. Андрєєвиє М. — дрібні червоно-бурі (до чорних), з одношаровим листям; зростають на скелях подушечками. Коробочка подовжений-яйцевидна, на псевдоніжці, розкривається 4—8 стулками, без устьіц, із споровим мішком у вигляді зведення над верхньою частиною колонки. Поширені високо в горах, а також в північних широтах. Брієвиє М. — найбільш обширна, всіляка група. Коробочка зазвичай на справжній ніжці, з устьіцамі, частіше після опадання кришечки відкривається вгорі широким отвором — гирлом. У багатьох по краю гирла утворюється 1—2 ряди виростов — зубців (званих простим або подвійним перистомом), які служать для розсіювання спор. Колонка, як правило, пронизує споровий мішок. (Цикл розвитку див.(дивися) Листяні мохи .)
М. поширені повсюдно. Поселяються всюди, окрім Морея, засолених грунтів і місцепроживань, прихованих під льодовиками або схильних до сильної ерозії. Антоцеротовиє і печінкові М. поширені в основному в країнах з тропічним або помірним вологим кліматом; лише небагато пристосувалися до зростання в сухих місцях. Листяні М. зростають майже повсюдно, проте найбільшого розвитку досягають на зволожених місцях, в лісах, тундрі; на болотах М. утворюють основну масу торф'яних покладів. Інтенсивно розвиваючись, М. сприяють заболочуванню грунтів, погіршують якість лугів і ін. з.-х.(сільськогосподарський) угідь. У зв'язку з антибіотичними властивостями деяких М. їх застосовують інколи як перев'язувальний матеріал. Використовують як підстилку для худоби (переважно сфагнові М.), в будівельній техніці для виготовлення плит і ін.
Літ.: Флора спорових рослин СРСР, т. 1, 3, М. — Л., 1952—54; Савіч-Любіцкая Л. І., Смирнова З. Н., Визначник сфагнових мохів СРСР, Л., 1968; їх же, Визначник лістостебельних мохів СРСР. Верхоплодниє мохи, Л., 1970; Smith G. M., Cryptogamic botany, 2ed., v. 2, N. Y., 1955; Schuster R. M., The Hepaticae and Anthocerotae of North America, v. 1, N. Y. — L., 1966; Parihar N. S., An introduction to embryophyta, 5 ed., v. 1, Allahabad, 1969.