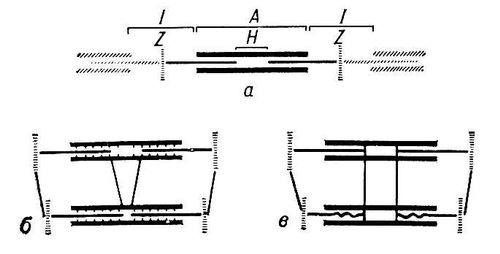
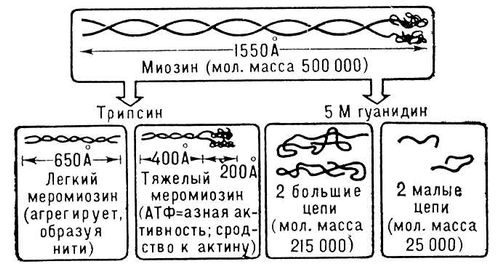
Мышечное сокращение, укорочение мышцы, в результате которого она производит механическую работу. М. с. обеспечивает способность животных и человека к произвольным движениям. Наиболее важная составная часть мышечной ткани — белки (16,5—20,9%), в том числе контрактильные, обусловливающие способность мышцы к сокращению. Значительный интерес представляют механоактивные миофибриллярные белки, изучение которых было начато В. Кюне (1864). Важные данные, характеризующие физико-химические и биохимические свойства механоактивных мышечных белков, были получены А. Я. Данилевским (1881—88). В 1-й половине 20 в. В. А. Энгельгардт и М. Н. Любимова (1939) установили, что основной контрактильный белок мышц — миозин — обладает аденозинтрифосфатазной активностью, а А. Сент-Дьёрдьи и Ф. Б. Штрауб показали (1942—43), что входящий в состав миофибрилл белок состоит в основном из 2 компонентов — миозина и актина. Взаимодействие этих фибриллярных белков и лежит в основе феномена сокращения самых различных контрактильных органелл и органов движения (см. Мышцы). Периодические изменение физического состояния мышечных белков, обусловливающее возможность попеременного сокращения и расслабления мышц и выполнения ими механической работы, очевидно, связано с определёнными дающими энергию биохимическими процессами. Энгельгардтом и Любимовой (1939—42) было обнаружено, что особым образом приготовленные из миозина нити при взаимодействии с раствором АТФ резко изменяют свои механические свойства (эластичность и растяжимость). Одновременно происходит расщепление АТФ с образованием АДФ и неорганического фосфата. Это открытие заложило фундамент для нового направления в биохимии — механохимии М. с. В дальнейшем Сент-Дьёрдьи и Штрауб показали, что истинным сократительным белком является не миозин, а его комплекс с актином — актомиозин. Сокращению при взаимодействии с АТФ подвергаются как вымоченные в воде или 50%-ном глицерине мышечные волокна, так и нити, приготовленные из актомиозиновых гелей (синерезис геля). Эти опыты подтверждают, что энергия, необходимая для сокращения мышц, освобождается в результате взаимодействия актомиозина с АТФ с расщеплением последнего на АДФ и H3PO4. При этом освобождается большое количество энергии (8—10 ккал, или 33,5—41,9 кдж, на 1 моль АТФ). Однако истинный механизм этой реакции остаётся всё ещё неясным. Полагают, что терминальная фосфатная группировка АТФ при взаимодействии с актомиозином переносится на миозин без промежуточного образования тепла с образованием богатой энергией фосфорилированной формы актомиозина, способной к сокращению. Молекулярная масса миозина, определённая методом ультрацентрифугирования, близка к 500 тыс. Молекула миозина может быть расщеплена без разрыва ковалентных связей на более мелкие субъединицы (рис. 1): 2 «тяжёлые» полипептидные цепочки с молекулярной массой свыше 210 000 и 2 (по др. данным, 3) коротких («лёгких») полипептида с молекулярной массой около 20 000 каждый. По данным электронной микроскопии, молекула миозина состоит как бы из 2 частей — утолщённой «головки» и длинного «хвоста». Общая длина молекулы — около 1600 А. Большое число соответственно расположенных в пространстве макромолекул миозина образует в поперечнополосатом волокне толстые (миозиновые) нити. В образовании поперечных мостиков между толстыми (миозиновыми) и тонкими (актиновыми) нитями непосредственно участвуют, по-видимому, «головки» миозиновых молекул. Молекулярная масса мономера актина близка к 46 000 (ранее принималась около 70 000). Установлена и его первичная структура: число, природа и последовательность включения в полипептидную цепь аминокислотных остатков. Молекулы фибриллярного актина (Ф-актина) образованы 2 спиральными цепочками, состоящими из множества бусинок — молекул глобулярного актина (мономера актина, или Г-актина). В саркомерах поперечнополосатого волокна нити Ф-актина пространственно отграничены от нитей миозина. Взаимодействие систем двух типов нитей осуществляется за счёт энергии, освобождающейся при расщеплении АТФ в присутствии ионов Ca2+ (рис. 2). Т. к. при работе мышц АТФ постоянно потребляется, для длительного осуществления двухфазной мышечной деятельности необходимо непрерывное восстановление АТФ — её ресинтез. Ресинтез АТФ из АДФ и H3PO4 сопряжён с рядом дающих энергию превращений. Важнейшие из них: 1) перенос фосфатной группы с фосфокреатина (КрФ) на АДФ. Эта реакция обеспечивает быстрый, происходящий уже во время сокращения мышцы, ресинтез АТФ за счёт потребления креатинфосфата; 2) гликогенолиз, или гликолиз (расщепление гликогена или глюкозы с образованием молочной кислоты); 3) тканевое дыхание (образование АТФ в митохондриях мышечных волокон за счёт энергии окисления главным образом углеводов, жирных кислот и ненасыщенных фосфолипидов). Некоторое количество АТФ может образовываться также в результате миокиназной реакции из АДФ: 2 АДФ >< АМФ + АТФ. Фосфорилирование креатина за счёт АТФ с образованием КрФ осуществляется в процессе гликолиза и тканевого дыхания. Ре-синтез КрФ и гликогена происходит главным образом в фазе отдыха после расслабления мышцы. Скелетная мышца, находящаяся в анаэробных условиях или в условиях кислородного голодания (гипоксии), способна к выполнению некоторого количества работы. Однако утомление в этих случаях наступает значительно раньше, чем в присутствии кислорода, и сопровождается накоплением в мышце молочной кислоты.
А. В. Палладиным, Д. Л. Фердманом, Н. Н. Яковлевым и др. получены данные о биохимической сущности тренировки мышц. С. Е. Севериным продемонстрирована способность дипептидов (карнозина, анзерина) восстанавливать работоспособность утомлённых мышц и влиять на передачу нервных импульсов с нерва на мышцу.
После сокращения, вызванного раздражением с нерва или электрическим током, мышца вскоре переходит в расслабленное состояние, хотя содержание АТФ в мышечных волокнах почти не меняется. Установлено, что миофибриллы обладают способностью взаимодействовать с АТФ и сокращаться в её присутствии лишь при наличии в среде ионов Ca2+. Наибольшая сократительная активность наблюдается при концентрации Ca2+ около 10-6—10-5моль. При понижении содержания Ca2+ до 10-7моль или ниже мышечные волокна теряют способность к укорочению и развитию напряжения (тянущей силы) в присутствии АТФ. По современным представлениям, в покоящейся мышце концентрация ионов Ca2+ поддерживается ниже этой пороговой величины вследствие их связывания структурами (трубочками и пузырьками) саркоплазматической сети. Связывание — это не простая адсорбция, а активный физиологический процесс, осуществляемый за счёт энергии, освобождающейся при расщеплении АТФ в присутствии ионов Mg. Этот механизм получил название Са-насоса (по аналогии с «натриевым насосом»). Т. о., пребывание живой мышцы (при наличии в ней достаточного количества АТФ) в расслабленном состоянии — результат снижения под действием Са-насоса концентрации ионов Ca2+ в среде, окружающей миофибриллы, ниже предела, при котором ещё возможно проявление АТФ-азной активности и сократимости актомиозиновых структур волокна. Сокращение волокна при раздражении с нерва (или электрическим током) — результат внезапного изменения проницаемости и, как следствие, выхода из цистерн и трубочек саркоплазматической сети и так называемой Т-системы ионов Ca2+ в межфибриллярное пространство. Поперечные трубочки Т-системы, расположенные на уровне Z-дисков и содержащие Ca2+, сообщаются с поверхностной мембраной волокна; поэтому волна деполяризации быстро распространяется по системе трубочек и достигает глубоко расположенных участков волокна. После затухания нервного импульса в результате действия Са-насоса концентрация Ca2+ в межфибриллярном пространстве быстро снижается до пороговой величины и мышца переходит в исходное расслабленное состояние, пока новый импульс не вызовет повторение всего цикла. Потерю актомиозином способности расщеплять АТФ и сокращаться при снижении концентрации ионов Ca2+ ниже 10-7моль связывают с присутствием в контрактильной системе особого белка — тропонина. При его отсутствии актомиозин реагирует in vitro (в пробирке) с АТФ практически и в отсутствии Ca2+. В физиологических условиях (in vivo) тропонин — постоянный компонент контрактильной системы мышечного волокна (тропонин-тропомиозиновый комплекс).
Двухфазный механизм М. с. не исчерпывается изложенными представлениями. У некоторых насекомых (жуки, пчёлы, мухи, комары и др.) частота сокращений мышц крыльев много выше частоты поступающих нервных импульсов. Эти мышцы подчинены не нейрогенному, а миогенному ритму. Они могут совершать колебания (осциллировать) несколько сот раз в 1 сек. Осцилляция этих мышц не связана с изменением концентрации Ca2+ в саркоплазме мышечных волокон. Возможность автоматической двухфазной деятельности клеточных органелл движения в присутствии АТФ можно наблюдать на клеточных моделях — сперматозоидах, мерцательном эпителии, ундулирующих мембранах трипаносом и др. Осцилляция органелл движения происходит с обычной для данного вида клеток скоростью при постоянной концентрации Ca и продолжается до тех пор, пока в растворе сохраняется известный избыток АТФ. Механизм такой осцилляции органелл движения, а также миофибрилл, по-видимому, может быть понят лишь исходя из существования взаимосвязи между ферментативной активностью (способностью расщеплять АТФ) и состоянием (конформацией) макромолекул сократительного субстрата.