Зародыш, 1) у животных и человека З., или эмбрион, — организм в раннем периоде развития — от оплодотворённого яйца (зиготы) до появления способности к самостоятельному питанию и активному передвижению. Развитие З. происходит, как правило, в яйцевых оболочках или специальных органах материнского организма. См. Зародышевое развитие, Зародышевые оболочки.
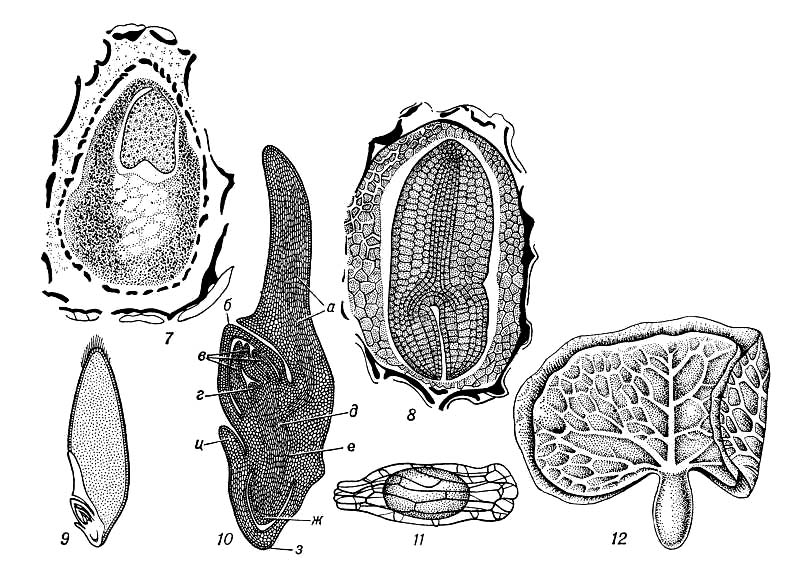
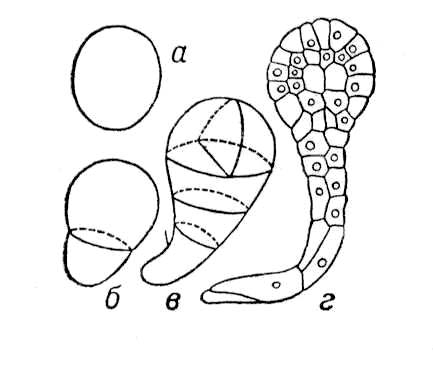
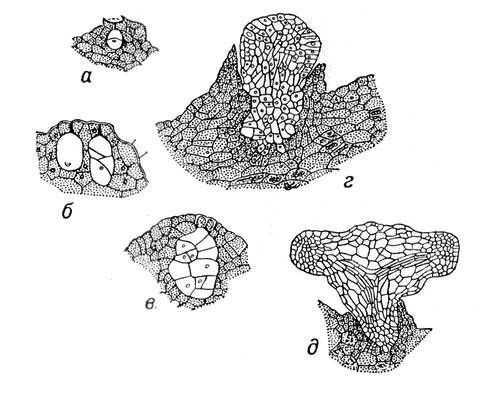
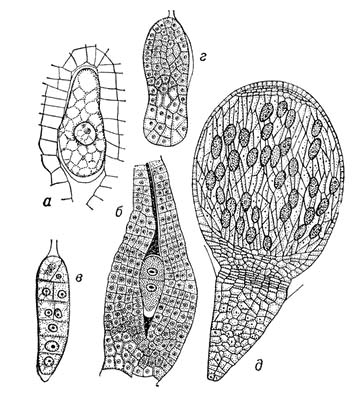
2) У растений З., или эмбрио, — зачаток новой особи, развивающийся из зиготы, которая возникает в результате оплодотворения яйцеклетки зародышевого мешка или из клеток нуцеллуса или интегумента (в случае апомиксиса). Наиболее простой тип эмбрионального развития у зелёных водорослей, например у спирогиры, зигота которой не образует спорофитной нити, а сразу приступает к мейозу. У бурых водорослей, например у фукуса, зигота прикрепляется к субстрату, делится, после чего образует эмбриональное тело с точкой роста таллома (Рис.1). У мохообразных З. прорастает на материнском организме — гаметофите, за счёт которого он питается, затем образует спорогенную ткань, где формируются споры (Рис.2). Такой тип З. не встречается среди др. групп высших растений. У древнейших высших растений — псилотовых — развитие З. протекает в ткани гаметофита, где происходит ряд последовательных делений зиготы. Из клеток нижней части формируется ножка, внедряющаяся в ткань гаметофита, в верхней — закладывается точка роста побега с двумя меристематическими бугорками (см. Меристема). З. здесь — поляризованная структура, один конец которой образует наземный побег, другой — корневище (Рис.3). У плауновидных З. имеет два симметричных бугорка будущих зародышевых листочков, между которыми находится точка роста в виде верхушечной клетки (Рис.4). Развитие З. у хвощей характеризуется правильным планом сегментации в виде 4 зон, дающих начало побегу, корню, первому листу и ножке (Рис.5). У папоротниковидных в первых 4 клетках, возникших в результате двух первых делении зиготы, определено место закладки верхушки побега, первого листа, корня и ножки (Рис.6). У голосеменных начальный цикл развития З. характеризуется свободноядерной, или ценоцитной, стадией (у саговников, например, свободных ядер 1024), которая сменяется образованием клеток; затем в формирующемся З. происходит закладка точек роста побега, корня и семядольных бугорков (у некоторых видов сосны — до 18) (Рис.7). У большинства покрытосеменных зигота делится на две клетки: апикальную, дающую начало собственно З., п базальную — подвеску, или суспензору.
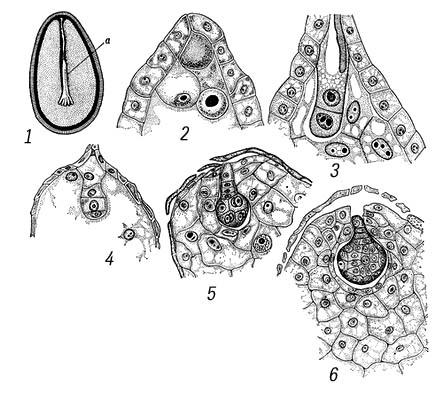
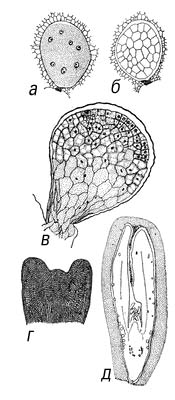
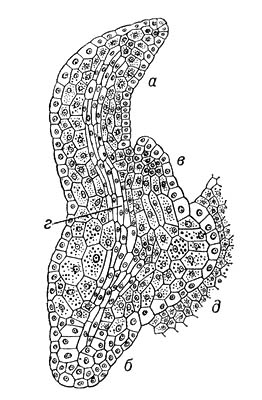
В эмбриогенезе различают 2 фазы развития: предзародышевую (от зиготы до образования эмбриодермы) и зародышевую (закладка основных структур: точки роста побега, семядоли, гипокотиля и центрального зародышевого корешка). На основе первых этапов деления предзародыша были созданы классификации эмбриональных типов, используемые систематиками при решении вопросов эволюции и филогении растений. Предзародышевая стадия развития двудольных и однодольных одинакова. На более поздних стадиях у однодольных апикальная зона проэмбрио даёт начало только 1 семядольному бугорку; у двудольных таких бугорков может возникнуть 2, а у некоторых видов 3 и даже больше, что приводит к образованию многосемядольности. У покрытосеменных растений развитие З., в отличие от голосеменных, сопровождается образованием клеточных перегородок (Рис.8,9). Лишь у пионов обнаружен новый тип эмбриогенеза, при котором в зиготе вначале возникают свободные ядра, а затем такая многоядерная структура становится клеточной и на её периферии возникают проэмбриональные бугорки; из них в собственно З. развивается только один. По наличию или отсутствию у З. покрытосеменных зелёного пигмента — хлорофилла — они делятся на 2 группы: хлороэмбриофиты (бобовые, крестоцветные и др.) илейкоэмбриофиты (лютиковые, злаковые и др.). В процессе эволюции среди некоторых групп покрытосеменных, например у растений паразитов (заразиховые) и сапрофитов (орхидные), наблюдается -редукция З. Источником питания З. служит эндосперм, который у одних видов (бобовые) поглощается целиком растущим З., у др. (злаки) — сохраняется. Иногда З. в семени окружен не только эндоспермом, но и др. запасной тканью — периспермом, развивающимся из клеток нуцеллуса (свёкла, чёрный перец и др.).
Лит.: Баранов П. А., История эмбриологии растений в связи с развитием представлений о зарождении организмов, М.— Л., 1955; Яковлев М. С., Эмбриогенез и его значение для филогении растений, М. — Л., 1960; Поддубная-Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Тахтаджян А. Л., Основы эволюционной морфологии покрытосеменных, М. — Л., 1964.