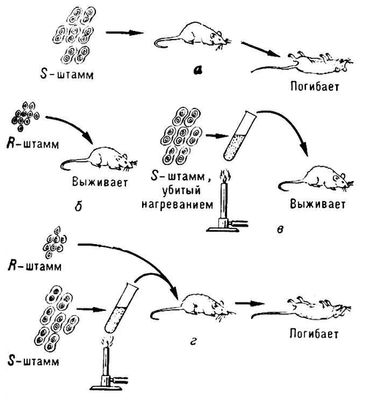
Трансформація в генетиці, внесення до клітки генетичної інформації за допомогою ізольованої дезоксирибонуклеїнової кислоти (ДНК). Т. приводить до появи в трансформованої клітки (трансформанта) і її потомства нових ознак, характерних для об'єкту — джерела ДНК(дезоксирибонуклеїнова кислота). Явище Т. було відкрите в 1928 англійським ученим Ф. Грифітом що спостерігав успадковане відновлення синтезу капсульного полісахариду в пневмококів при зараженні мишей сумішшю убитих нагріванням капсулірованних бактерій і кліток, позбавлених капсули. Організм миші в цих експериментах грав роль своєрідного детектора, оскільки придбання капсульного полісахариду повідомляло клітки, позбавлені капсули, здатність викликати смертельний для тварини інфекційний процес (див. схему ). У подальших експериментах було встановлено, що Т. має місце і у тому випадку, коли замість убитих кліток до позбавлених капсули пневмококів додавали екстракт із зруйнованих капсулірованних бактерій. У 1944 О. Ейвері із співробітниками (США) встановив, що чинником, що забезпечує Т., є молекули ДНК(дезоксирибонуклеїнова кислота). Ця робота — перше дослідження, що довело роль ДНК(дезоксирибонуклеїнова кислота) як носія спадкової інформації.
Окрім пневмококів Т. виявлена і вивчена на деяких інших бактеріях. Використання в експериментах генетичних ознак (наприклад, стійкість до дії клітинних отрут, потреба в певних чинниках зростання), що легко враховуються, а також вживання ДНК(дезоксирибонуклеїнова кислота) з радіоізотопною міткою дозволили дати Т. кількісну оцінку. Т. у бактерій розглядають як складний процес, що включає наступні стадії: фіксація молекул ДНК(дезоксирибонуклеїнова кислота) кліткою-реципієнтом; проникнення ДНК(дезоксирибонуклеїнова кислота) всередину клітки; включення фрагментів трансформуючою ДНК(дезоксирибонуклеїнова кислота) в хромосому клітки-господаря; формування «чистих» трансформованих варіантів. Фіксація ДНК(дезоксирибонуклеїнова кислота) відбувається на особливих ділянках клітинної поверхні (рецепторах), число яких обмежене. Пов'язана з рецепторами ДНК(дезоксирибонуклеїнова кислота) зберігає чутливість до дії доданого в середу ферменту Дезоксирибонуклеази, що викликає її розпад. Проте, опісля дуже короткий термін (в межах 1 мін ) після фіксації, частина ДНК(дезоксирибонуклеїнова кислота) проникає в клітку. Бактерійні клітки одного і того ж штаму різко розрізняються по проникності для ДНК(дезоксирибонуклеїнова кислота). Клітки даної бактерійної популяції, здатні включати чужорідну ДНК(дезоксирибонуклеїнова кислота), називаються компетентними. Число компетентних кліток в популяції трохи і залежить від генетичних особливостей бактерій і фази зростання бактерійної культури. Розвиток компетенції пов'язують з синтезом особливого білка, що забезпечує проникнення ДНК(дезоксирибонуклеїнова кислота) в клітку.
Середні розміри фрагментів ДНК(дезоксирибонуклеїнова кислота), проникаючої в клітку, складають 5×10 6 дальтон. Оскільки в компетентну клітку може одночасно проникнути ряд таких фрагментів, сумарна величина поглиненої ДНК(дезоксирибонуклеїнова кислота) може приблизно дорівнювати розмірам хромосоми клітки-господаря. Після проникнення в клітку двониткової ДНК(дезоксирибонуклеїнова кислота) одна нитка розпадається до моно- і олігонуклеотідов, друга, — вбудовується в хромосому клітки-господаря шляхом її розривів і возз'єднань. Подальша реплікація такою гібридною структури приводить до вищепленію «чистих» клонів трансформантов, в потомстві яких закріплена ознака, що кодується ДНК(дезоксирибонуклеїнова кислота), що включилася.
Вживання Т. дозволило провести генетичний аналіз бактерій, в яких не описано інших форм генетичного обміну (кон'югації, трансдукції ). Крім того, Т. — зручний метод для з'ясування впливів на біологічну активність ДНК(дезоксирибонуклеїнова кислота) фізичних або хімічних змін її структури. Розробка методу Т. в кишкової палички дозволила використовувати для Т. не лише фрагменти бактерійної хромосоми, але і ДНК(дезоксирибонуклеїнова кислота) бактерійних плазмід і бактеріофагів . Цей метод широко використовується для внесення до клітки гібридної ДНК(дезоксирибонуклеїнова кислота) в дослідженнях по так званій генній інженерії.
Є повідомлення про відтворення Т. на клітках вищих організмів. Проте в цьому випадку процес Т. вивчений недостатньо.
Літ.: Хейс В., Генетика бактерій і бактеріофагів, пер.(переведення) з англ.(англійський), М., 1965; Прозоров А. А., Генетична трансформація у мікроорганізмів, М., 1966; Браун Ст, Генетика бактерій, пер.(переведення) з англ.(англійський), М., 1968; Бреслер С. Е., Молекулярна біологія, Л., 1973; Стент Р., Молекулярна генетика, пер.(переведення) з англ.(англійський), М., 1974, гл.(глав) 7.