Стелярная теорія, вчення про принципи будови і взаємини між типами стели (центрального циліндра ) у вищих рослин. Велику частину стели складають провідні тканини : ксилема (деревина) і флоема (луб), по-різному розташовані в різних типах стели. Провідні тканини зазвичай оточені перициклом, що складається з механічних і паренхімних кліток. Довкола стели знаходиться первинна кора.
Вчення про стеле створене французькими ботаніками Ф. Ван Тігемом і А. Дуліо (1886), яким належить і перша класифікація типів стели. Подальший розвиток С. т. отримала в роботах англійського ботаніка Г. Бребнера (1902), американського, — Е. Джефрі (1903, 1917), що значно удосконалили класифікацію типів стели, а потім в працях німецького ученого В. Циммерманна і радянських — До. І. Мейєра, А. Л. Тахтаджяна, що відзначили зміни стели в онтогенезі і філогенезі рослин.
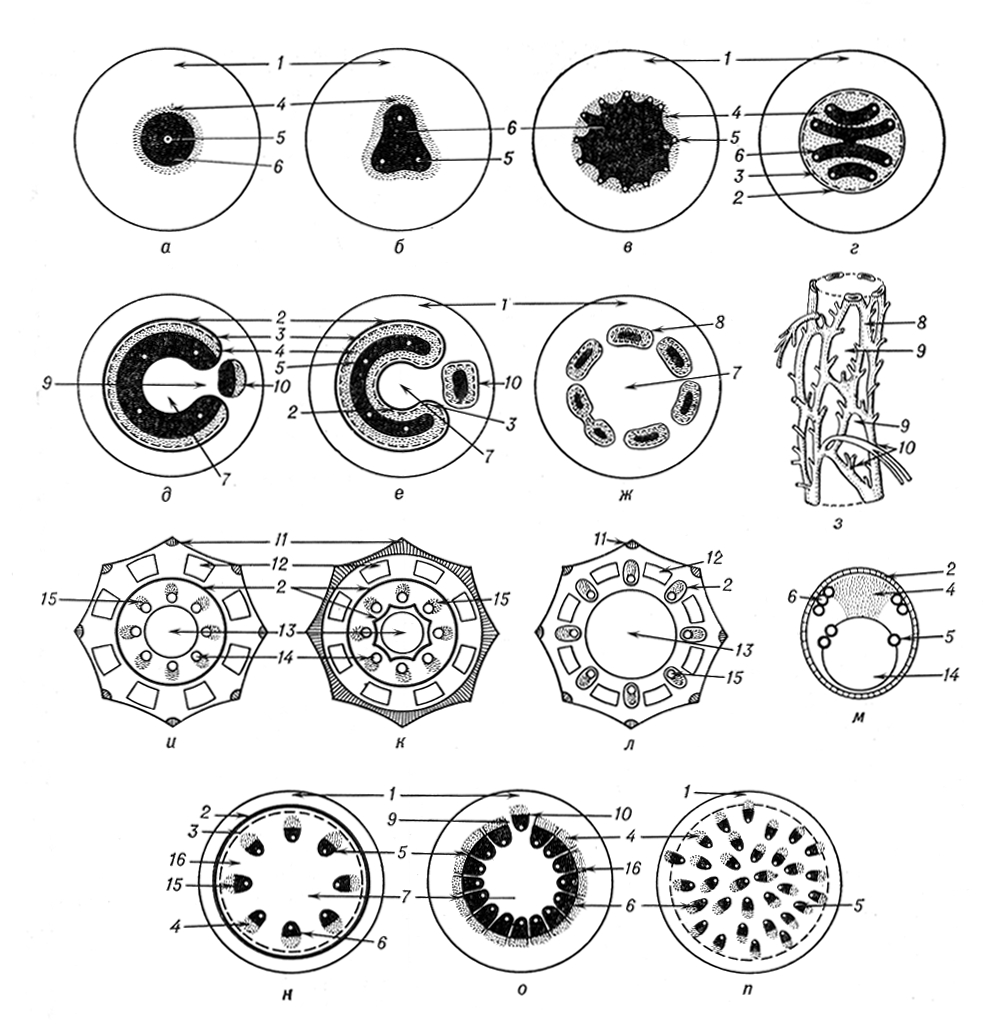
Вихідним типом стели, характерним для прадавніх вищих рослин, — псилофітов (рінія), вважають протостелу, що має вигляд центр, тяжа, у внутрішній частині якого розташована ксилема, оточена нерізко відмежованою від первинної кори флоемою. Вдосконалення структури стели в еволюції рослин, на думку англійського ботаніка Ф. Боуера, йшло по шляху створення найбільш раціонального співвідношення між об'ємом і поверхнею провідних тканин, що досягалося змінами контурів стели і привело до розчленовування її на окремі тяжі. Велику роль в еволюції стели зіграли процеси медулляциі, тобто формування серцевини, і віталізациі провідних тканин — появи в них живих паренхімних клітин. Розвиток стелярной структури рослин супроводилося також диференціацією прокамбію на протоксилему і метаксилему, протофлоему і метафлоему, змінами характеру заставляння ксилеми від мезархного до екзархному і ендархному. При мезархному заставлянні перші елементи ксилеми диференціюються в центрі прокамбіального тяжа, все подальші — в радіальних напрямах. Для екзархной ксилеми характерне периферичне заставляння перших елементів і доцентровий розвиток подальших. У ендархной ксилемі перші елементи формуються з внутрішньої частини прокамбію, а наступні розвиваються відцентровий. Велике значення мали розчленовування стели на стеблові і листові пучки, що входять в стебло з аркуша, посилення зв'язку між ними, формування ендодерми як бар'єру, що запобігає втраті вологи і що затримує в стеле продукти асиміляції. В процесі історичного розвитку наземних рослин відбувалося, як правило, збільшення розмірів стели.
В різних напрямах еволюції рослин виникали різні типи структурної організації стели. Так, зміни контурів ксилеми зумовили перетворення протостели в актіностелу і плектостелу. Актіностела, характерна для псилофітов (астероксилон), а з сучасних рослин — для псилота, має лопатеві контури поперечного перетину екзархной ксилеми.
В плектостеле, звичайною для плавунів, екзархная ксилема розітнута на стрічкоподібні тяжі. Система провідних тканин, створюючих трубчастий циліндр, що оточує паренхімну серцевину, характерна для сифоностели. В папоротей сифоностела буває трьох типів: ектофлойная (соленоксилія), амфіфлойная (соленостела) і діктіостела. Соленоксилія утворилася, мабуть, з актіностели при втягуванні всередину відрогів ксилеми, зрощенні ділянок флоеми в суцільне кільце і розвитку паренхімної серцевини, клітки якої виникли з трахеїдів, що втратили здатність проводити воду і що поділилися поперечними перегородками. У соленоксилії ксилема зовні оточена флоемою, перициклом і ендодермою (наприклад, в хельмінтостахиса). У соленостеле є не лише зовнішні, але і внутрішні флоема, перицикл і ендодерма (наприклад, в марсилії). Дослідження К. І. Мейера по розвитку провідної системи папоротей показали можливість утворення соленостели з соленоксилії. В результаті сильного розтину амфіфлойной сифоностели у зв'язку з появою багаточисельних аркушевих проривів, заповнених паренхімою, виникла діктіостела, що має вигляд сітчастого циліндра, т.к. составляющие її провідні тканини утворюють тяжі, що переплітаються (мерістели). На поперечному зрізі стебла мерістели розташовані кільцем довкола серцевини. Вони побудовані за типом концентричних амфікрібральних пучків, в яких флоема розташована довкола ксилеми; флоема оточена перициклом і ендодермою.
Вікові зміни стели в папоротей (мараттія орляк, матонія), що виражаються у формуванні усередині однієї стели другої, а потім і третьою, привели до поліциклії. Перехід від протостели до полістелії зумовив, на думку. Ст Циммерманна, формування еустели, в якій кожна протостела перетворилася на колатеральний пучок. Деякі ботаніки вважають, що еустела могла сформуватися з ектофлойной сифоностели, розчленовування якої на окремі пучки було викликане утворенням серцевинних променів. В хвощів еустела представлена закритими колатеральними пучками, що розташованими довкола центральної повітряної порожнини і з'єднуються у вузлах; на місці ксилеми, що рано руйнується, в пучку утворюється карінальний канал (водоносна порожнина). Цей різновид стели називають артростелой В різних видів хвощів артростели характеризуються різним розташуванням ендодерми (див. мал.(малюнок), позиції і, до, л) . Дослідження ранніх етапів розвитку провідної системи хвощів дають можливість передбачати що артростела утворилася з актіностели або з сифоностели унаслідок розщеплювання їх на окремі пучки. Для еустели насінних рослин характерне посилення ролі листових пучків і наявність тісних контактів між стебловими і листовими пучками. Еустела дводольних рослин представлена системою відкритих колатеральних або біколлатеральних пучків з ендархной первинною ксилемою; пучки розділені паренхімними серцевинними променями, що пересікають стелу в радіальному напрямі. В багатьох трав'янистих рослин серцевинні промені широкі; в деревинних — вузькі, інколи однорядні, перициклу і ендодерми немає.
Потужний розвиток провідних пучків листя, яке, увійшовши до стебла, розташовується по всьому поперечному перетину стебла, а також редукція стеблових пучків привели до перетворення еустели в атактостелу однодольних рослин, що втратила здібність до вторинного потовщення. Провідні пучки в атактостеле колатеральні або концентричні амфівазальниє (ксилема в них оточує флоему).
Виявляючи істотні відмінності між основними відділами вищих рослин по архітектоніці їх провідної системи, С. т. має велике значення не лише для анатомії рослин, але і для пізнання філогенезу рослин в цілому.
Літ.: Раздорський Ст Ф., Анатомія рослин, М-код,, 1949; Чистякова О. Н., Історія розвитку провідної системи в хвощів, «Уч. зап.(західний) Московського міського педагогічного інституту», 1955, т. 29. ст 3; Тахтаджян А. Л., Вищі рослини, t. i, М. — Л., 1956; Мейєр До. І., Морфогенія вищих рослин, [М.], 1958: Zimmermann W., Die Pliylogenie der Pflanzen, 2 Aufl., Stuttg., 1959.