Плавники, органи руху водних тварин. Серед безхребетних П. мають пелагічні форми брюхоногих і головоногих молюсків і щетінко-щелепні. У брюхоногих молюсків П. є видозміненою ногою, в головоногих — бічні складки шкіри. Для щетінкочелюстних характерні бічні і хвостовий П., утворені складками шкіри. Серед сучасних хребетних П. мають круглоротиє, риби, деякі земноводні і ссавці. В круглоротих — лише непарні П.: передній і задній спинний (у міног) і хвостовий.
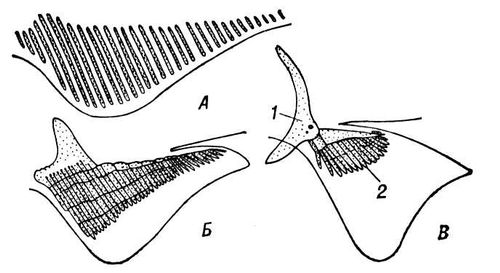
У риб розрізняють парні і непарні П. Парниє представлені передніми (грудними) і задніми (черевними). У деяких риб, наприклад у тріскових і морських собачок, черевні П. інколи розташовані попереду грудних. Скелет парних П. складається з хрящових або кісткових променів, які прічленяются до скелета поясів кінцівок ( мал. 1 ). Основна функція парних П.— напрям руху риби у вертикальній плоскості (кермо глибини). В ряду риб парні П. виконують функції органів активного плавання або служать для планерування в повітрі (у летких риб), повзання по дну або пересування по суші (у риб, що періодично виходять з води, наприклад у представників тропічного роду Periophtalmus, які при допомозі грудних П. можуть навіть влізати на дерева). Скелет непарних П.— спинного (часто розділеного на 2, а інколи на 3 частини), заднепроходного (інколи розділеного на 2 частини) і хвостового — складається з хрящових або кісткових променів, лежачих між бічними м'язами тіла ( мал. 2 ). Скелетні промені хвостового П. пов'язані із заднім кінцем хребта (у деяких риб вони замінюються остистими відростками хребців).
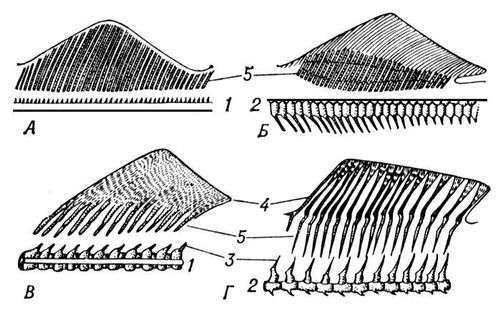
Периферичні частини П. підтримуються тонкими променями з рогоподібної або кісткової тканини. У колючоперих риб передні з цих променів товщають і утворюють тверді колючки, інколи пов'язані з отруйними залозами. До підстави цих променів прикріпляються м'язи, що розтягують лопать П. Спінной і заднепроходний П. служать для регулювання напряму руху риби, але інколи вони можуть бути і органами поступальної ходи або виконувати додаткові функції (наприклад, залучення видобутку). Хвостовий П., сильно що варіює формою у різних риб, - основний орган руху.
В процесі еволюції хребетних П. риб виникли, ймовірно, з суцільної шкірної складки, яка проходіла уздовж спини тварини, огинала задній кінець його тіла і продовжувалася на черевну сторону до заднепроходного отвору, потім розділялася на дві бічні складки, що тривали до зябрових щілин; таке положення плавникових складок в сучасного примітивного хордового — ланцетника . Можна передбачити, що в процесі еволюції тварин в деяких місцях таких складок утворилися скелетні елементи і в проміжках складки зникли, що привело до виникнення непарних П. в круглоротих і риб і парних — у риб. На користь цього говорить знаходження бічних складок або отрути Шипа в прадавніх хребетних (деякі безщелепні, акантодії) і те, що у сучасних риб парні П. мають велику протяжність на ранніх стадіях розвитку, чим в дорослому стані. Серед земноводних непарні П. у вигляді шкірної складки, позбавленої скелета, є як постійні або тимчасові утворення у більшості личинок, що живуть у воді, а також у дорослих хвостатих і личинок безхвостих земноводних. Серед ссавців П. є в тих, що перейшли повторно до водного способу життя китоподібних і бузкових. Непарні П. китоподібних (вертикальний спинний і горизонтальний хвостовий) і бузкових (горизонтальний хвостовий) не мають скелета; це вторинні утворення, не гомологічні (див. Гомологія ) непарним П. риб. Парні П. китоподібних і бузкових, представлених лише передніми П. (задні зредуковані), мають внутрішній скелет і гомологични переднім кінцівкам всіх ін. хребетних.
Літ. Керівництво по зоології, т. 2, М.— Л., 1940; Шмальгаузен І. І., Основи порівняльної анатомії хребетних тварин, 4 видавництва, М., 1947; Суворов Е. До., Основи іхтіології, 2 видавництва, М., 1947; Догель Ст А., Зоологія безхребетних, 5 видавництво, М., 1959; Алєєв Ю. Р., Функціональні основи зовнішньої будови риби, М., 1963.